von Ana Catarina Ribeiro Carrão
Statistik und Sichtungsnachweis dieser Seite findet sich am Artikelende
| [1.] Arc/Fragment 037 01 - Diskussion Zuletzt bearbeitet: 2014-03-06 09:16:13 Singulus | Arc, BauernOpfer, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Ushio-Fukai 2006 |
|
|
|
| Untersuchte Arbeit: Seite: 37, Zeilen: 0-1 |
Quelle: Ushio-Fukai 2006 Seite(n): 231, Zeilen: 1ff |
|---|---|
| [In Fig. 10, Ushio-Fukai describes important ROS-dependent genes and proteins associated with anigiogenesis, like urokinase plasminogen activator (uPA), plasminogen activator inhibitor-1 (PAI-1) and metalloproteinases (MMPs).]
Fig. 10 - Role of ROS derived from NADPH oxidase in the induction of transcription factors and genes involved in angiogenesis. Ischemia/hypoxia stimulates VEGF induction and NADPH oxidase, thereby promoting activation of redox signalling events and induction of redox sensitive transcription factors and genes involved in angiogenesis. ROS derived from NADPH oxidase also upregulate mitochondrial SOD (MnSOD), increasing mitochondrial H2O2 production, which could represent a feed-forward mechanism by which ROS-triggered H2O2 formation plays an important role in angiogenesis101. 101. Tuttle JL, Sanders BM, Burkhart HM, Fath SW, Kerr KA, Watson WC, Herring BP, Dalsing MC, Unthank JL. Impaired collateral artery development in spontaneously hypertensive rats. Microcirculation. 2002;9:343-351. |
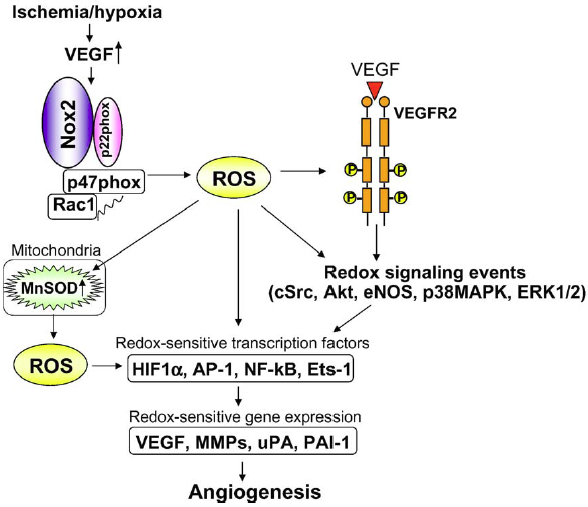
Fig. 4. Role of ROS derived from NADPH oxidase in induction of transcription factors and genes involved in angiogenesis. Ischemia/hypoxia stimulates VEGF induction and NADPH oxidase, thereby promoting activation of redox signaling events and induction of redox sensitive transcription factors and genes involved in angiogenesis. ROS derived from NADPH oxidase also upregulate MnSOD which increases mitochondrial H2O2 production, which could represent a feed-forward mechanism by which ROS-triggered H2O2 formation plays an important role in angiogenesis. |
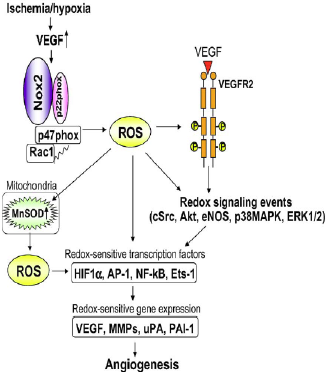
On the previous page it is mentioned that "In Fig. 10, Ushio-Fukai describes important ...". However, there is no reference to a publication by Usio-Fukai to be found in the list of references, and it is also not clear that the caption of figure 10 is also taken from that source. In the given source Tuttle et al. (2002) none of the copied material can be found. |
|
| [2.] Arc/Fragment 037 11 - Diskussion Zuletzt bearbeitet: 2014-02-26 00:43:54 Hindemith | Arc, Fragment, Gesichtet, Maulik et al 1998, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 37, Zeilen: 11-16 |
Quelle: Maulik et al 1998 Seite(n): 365, Zeilen: l.col: 33-40 |
|---|---|
| Tissue such as the myocardium can be adapted to ischemic stress by repeatedly subjecting it to short terms of reversible ischemia, each followed by another short duration of reperfusion108, 109. This phenomenon causes the production of oxidative stress, leading to the induction of gene expression, which is subsequently translated into the development of beneficial proteins responsible for the heart’s defence97.
97. Murphy E, Steenbergen C. Mechanisms Underlying Acute Protection From Cardiac Ischemia-Reperfusion Injury. Physiol. Rev. 2008;88:581-609. 108. Maulik N, Yoshida T, Zu YL, Sato M, Banerjee A, Das DK. Ischemic preconditioning triggers tyrosine kinase signaling: a potential role for MAPKAP kinase 2. Am J Physiol. 1998;275:H1857-1864. 109. Maulik N, Yoshida T, Engelman RM, Deaton D, Flack JE, 3rd, Rousou JA, Das DK. Ischemic preconditioning attenuates apoptotic cell death associated with ischemia/reperfusion. Mol Cell Biochem. 1998;186:139-145. |
Mammalian heart can be adapted to ischemia by repeatedly subjecting it to short-term reversible ischemia each followed by another short duration of reperfusion [1,2]. This phenomenon, known as ischemic adaptation, causes the production of oxidative stress leading to the induction of gene expression which is subsequently translated into the development of several stress-related proteins responsible for the heart's defense [3].
[1] Reimer, A., vander Heide, R.S. and Jennings, R.B. (1994) Ann. NY Acad. Sci. 723, 99-115. [2] Banerjee, A., Locke-Winter, C., Rogers, K.B., Mitchell, M.B., Bensard, D.D., Brew, E.C., Cairns, C.B. and Harken, A.H. (1993) Circ. Res. 73, 656-670. [3] Das, D.K., Maulik, N. and Moraru, I.I. (1995) J. Mol. Cell. Cardiol. 27, 181-193. |
The source is not given (the two papers by Maulik et al. given are different to the source here documented). The papers referenced under 97, 108 and 109 do not contain the copied text. |
|
Letzte Bearbeitung dieser Seite: durch Benutzer:Hindemith, Zeitstempel: 20140226004419