41 gesichtete, geschützte Fragmente: Plagiat
| [1.] Arc/Fragment 012 03 - Diskussion Bearbeitet: 25. February 2014, 21:58 Hindemith Erstellt: 22. February 2014, 13:41 (Graf Isolan) | Arc, Demetri and Griffin 1991, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 12, Zeilen: 3-11 |
Quelle: Demetri and Griffin 1991 Seite(n): 2791, Zeilen: left col. 1-14 |
|---|---|
| Granulocyte-colony stimulating factor (G-CSF) is a polypeptide growth factor that regulates the production of neutrophilic granulocytes. This physiological process serves as the foundation for critical host defence systems and occurs on a large scale in vivo. An adult of average size will produce approximately 120 billion granulocytes per day simply to replace normal losses1. This enormous production capacity may be increased by at least 10-fold under stress conditions such as infection. G-CSF plays a pivotal role in the basal regulation of neutrophil production as well as functioning as a primary regulatory factor controlling the neutrophil response to inflammatory stimuli. Also, G-CSF exhibits other biological activities and G-CSF-induced hematopoietic stem cell mobilization is widely used clinically for peripheral blood stem cell transplantation.
1. Basu S, Dunn A, Ward A. G-CSF: function and modes of action (Review). Int J Mol Med. 2002;10:3-10. |
GRANULOCYTE colony-stimulating factor (G-CSF) is a polypeptide growth factor that regulates the production of neutrophilic granulocytes. This physiologic process serves as the foundation for critical host defense systems and occurs on a large scale in vivo. An adult of average size will produce approximately 120 billion granulocytes per day simply to replace normal losses. This enormous production capacity may be increased by at least 10-fold under stress conditions such as infection. G-CSF is likely to play a role in the basal regulation of neutrophil production as well as to function as a primary regulatory factor controlling the neutrophil response to inflammatory stimuli. Further, G-CSF exhibits other biologic activities besides proliferative effects: [...] |
This passage is not recognizable as a citation. |
|
| [2.] Arc/Fragment 012 19 - Diskussion Bearbeitet: 26. February 2014, 21:57 Schumann Erstellt: 22. February 2014, 14:12 (Graf Isolan) | Arc, BauernOpfer, Demetri and Griffin 1991, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 12, Zeilen: 19-29 |
Quelle: Demetri and Griffin 1991 Seite(n): 2791, Zeilen: left col. 16-32 |
|---|---|
| G-CSF possesses unique and interesting characteristics among the family of hematopoietic growth factors. This chapter will summarize the current state of knowledge of the structure and function of G-CSF and its receptor.
I.1 Identification of G-CSF and its gene The identification of CSFs was made possible by cell culture assays for hematopoietic progenitor cells developed in the mid 1960s by Metcalf and his colleagues2. These in vitro systems showed that the survival, proliferation, and differentiation of immature hematopoietic cells were dependent on the continued presence of humoral factors, which was collectively termed “colony-stimulating activity” (CSA)3. Before the purification of individual factors, early sources of CSA [included media that was conditioned by stimulated cultures of normal blood or certain tumour cells.] 2. Bradley TR, Robinson W, Metcalf D. Colony production in vitro by normal polycythaemic and anaemic bone marrow. Nature. 1967;214:511. 3. Demetri GD, Griffin JD. Granulocyte colony-stimulating factor and its receptor. Blood. 1991;78:2791-2808. |
G-CSF possesses unique and interesting characteristics among the family of hematopoietic growth factors. This review will summarize the current state of knowledge of the structure and function of G-CSF and its receptor.
IDENTIFICATION OF G-CSF The identification of CSFs was made possible by the cell culture assays for hematopoietic progenitor cells, which were developed in the mid 1960s independently by Metcalf, Sachs, and their colleagues. These in vitro systems showed that the survival, proliferation, and differentiation of immature hematopoietic cells were dependent on the continued presence of humoral factors, which were collectively termed “colony-stimulating activity” (CSA). Before the purification of individual factors, early sources of CSA included media conditioned by culture with stimulated normal blood or splenic leukocytes, placenta, or certain tumor cells1-5. 1. Metcalf D: The granulocyte-macrophage colony-stimulating factors. Science 229:16, 1985 2. Metcalf D: The molecular control of cell division, differentiation commitment and maturation in haemopoietic cells. Nature 339:27, 1989 3. Sachs L The molecular control of blood cell development. Science 238:1374, 1987 4. Quesenberry P, Levitt L Hematopoietic stem cells. N Engl J Med 301:755,1979 5. Golde D, Cline M: Regulation of granulopoiesis. N Engl J Med 291:1388,1974 |
At the end of the page the source is given. Nevertheless nothing has been marked as a citation and it is not clear to the reader that even the end of the previous section is taken verbatim from the source. |
|
| [3.] Arc/Fragment 013 01 - Diskussion Bearbeitet: 26. February 2014, 22:03 Schumann Erstellt: 22. February 2014, 15:29 (Graf Isolan) | Arc, BauernOpfer, Demetri and Griffin 1991, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 13, Zeilen: 1-6, 8-34 |
Quelle: Demetri and Griffin 1991 Seite(n): 2791-2792, Zeilen: 2791:left col. 29-32.34-37.39-43 - right col. 1-2.4-29.33-36 - 2791:left col. 1-2.26-30.32-35 |
|---|---|
| [Before the purification of individual factors, early sources of CSA] included media that was conditioned by stimulated cultures of normal blood or certain tumour cells. It was initially unclear whether these complex mixtures contained both individual factors specific for proliferation and/or separate factors that would specifically induce differentiation. Further investigation showed that many of these biological activities were attributable to the simultaneous presence of multiple factors in the crude medium. [In 1978, Byrne et al.4 reported that medium that was conditioned by mouse heart tissue was found to contain CSFs which produce in vitro colonies of granulocytes and/or macrophages.] Purification of these CSFs proved difficult and for many factors, expression, cloning and production of recombinant proteins were required to completely define the unique biological properties of individual CSFs. G-CSF was probably first identified as having a distinct activity by Burgess and Metcalf5, not by its ability to stimulate proliferation, but rather by the capacity of postendotoxin-treated mouse serum to induce differentiation in a murine leukemic cell line. Therefore, Metcalf initially termed G-CSF a granulocyte-macrophage differentiation factor (GM-DF) and he noted it to be related to (or being the same as) the so-called macrophage- and granulocyte-inducing proteins, reported by Lotem et al.6. GM-DF was shown to be separate from GM-CSF, which had been partially purified in the late 1970s. This distinction was experimentally determined by the generation of neutralizing antiserum that could block the effects of GM-CSF, but which failed to block the activity of GM-DF. GM-DF was then shown to co-purify with a novel medium that selectively stimulated the formation of granulocytic colonies from normal hematopoietic progenitor cells in vitro7 and, after further purification, this factor was ultimately renamed G-CSF8. Nicola et al.8 described the biochemical characteristics of murine G-CSF in 1983, as a hydrophobic glycoprotein with an apparent molecular weight of 24-25 Kd, containing a neuraminic acid moiety and at least one internal disulfide bond that was necessary for its biological activity. After the identification of the murine G-CSF, a human molecule with analogous activities was discovered9. Both the proliferation- and differentiation-inducing activities of the murine and human G-CSF molecules cross species boundaries, in contrast to other hematopoietic growth factors such as GM-CSF or interleukin-3 (IL-3) which are active in a species-specific manner on the neutrophilic cell lineage3. In 1986, Nomura et al.10 finally described G-CSF as a molecule that specifically induces growth of the neutrophilic granulocyte lineage of cells.
Further understanding of the biological and biochemical properties of G-CSF was greatly facilitated by cloning of the gene encoding G-CSF and the production of [recombinant protein for study11, 12.] 3. Demetri GD, Griffin JD. Granulocyte colony-stimulating factor and its receptor. Blood. 1991;78:2791-2808. 4. Byrne PV, Heit W, Kubanek B. Stimulation of in vitro granulocyte--macrophage colony formation by mouse heart conditioned medium. Br J Haematol. 1978;40:197-204. 5. Burgess AW, Metcalf D. Characterization of a serum factor stimulating the differentiation of myelomonocytic leukemic cells. Int J Cancer. 1980;26:647-654. 6. Lotem J, Lipton JH, Sachs L. Separation of different molecular forms of macrophage- and granulocyte-inducing proteins for normal and leukemic myeloid cells. Int J Cancer. 1980;25:763-771. 7. Metcalf D. Clonal extinction of myelomonocytic leukemic cells by serum from mice injected with endotoxin. Int J Cancer. 1980;25:225-233. 8. Nicola NA, Metcalf D, Matsumoto M, Johnson GR. Purification of a factor inducing differentiation in murine myelomonocytic leukemia cells. Identification as granulocyte colony-stimulating factor. J Biol Chem. 1983;258:9017-9023. 9. Nicola NA, Begley CG, Metcalf D. Identification of the human analogue of a regulator that induces differentiation in murine leukaemic cells. Nature. 1985;314:625-628. 10. Nomura H, Imazeki I, Oheda M, Kubota N, Tamura M, Ono M, Ueyama Y, Asano S. Purification and characterization of human granulocyte colonystimulating factor (G-CSF). Embo J. 1986;5:871-876. 11. Nagata S, Tsuchiya M, Asano S, Kaziro Y, Yamazaki T, Yamamoto O, Hirata Y, Kubota N, Oheda M, Nomura H, Ono M. Molecular cloning and expression of cDNA for human granulocyte colony-stimulating factor. Nature. 1986;319:415-418. 12. Souza LM, Boone TC, Gabrilove J, Lai PH, Zsebo KM, Murdock DC, Chazin VR, Bruszewski J, Lu H, Chen KK, et al. Recombinant human granulocyte colony-stimulating factor: effects on normal and leukemic myeloid cells. Science. 1986;232:61-65. |
[Page 2791]
Before the purification of individual factors, early sources of CSA included media conditioned by culture with stimulated normal blood or splenic leukocytes, placenta, or certain tumor cells1-5. [...] It was initially unclear whether the complex mixtures termed CSA contained individual factors specific for proliferation and separate factors that specifically induced differentiation. [...] Further investigation showed that many of these biologic activities were attributable to the simultaneous presence of multiple factors in the crude CSA. Purification of the CSFs proved difficult, and, for many factors, expression cloning and production of recombinant protein were required to completely define the unique biologic properties of individual CSFs. [...] G-CSF was probably first identified as a distinct activity by Burgess and Metcalf, not by its ability to stimulate proliferation, but rather by the capacity of postendotoxin-treated mouse serum or conditioned media to induce differentiation of a murine myelomonocytic leukemia cell line, the differentiation-responsive (D+) subline of WEHI3B cells6,7. Therefore, G-CSF was initiaIly termed a granulocyte-macrophage differentiation factor (GM-DF) by Metcalf’ and was noted to be related to (or the same as) a differentiating activity named MGI-1G by Lotem et al.8 G-CSF was shown to be separate from GM-CSF, which had been partially purified in the late 1970s. This distinction was experimentally determined by the generation of neutralizing antisera that could block the effects of GM-CSF but which failed to block the activity of GM-DF. GM-DF was then shown to copurify with a novel activity that selectively stimulated the formation of granulocytic colonies by normal hematopoietic progenitor cells in vitro,’ and after further purification, this factor was ultimately renamed G-CSF.9 Nicola et al9 described the biochemical characteristics of murine G-CSF in 1983 as a hydrophobic glycoprotein with an apparent molecular weight of 24 or 25 Kd, containing a neuraminic acid moiety and at least one internal disulfide bond necessary for biologic activity. After the identification of the murine G-CSF, a human molecule with analogous activities was discovered. [...] Both the proliferation- and differentiation-inducing activities of the murine and human G-CSF molecules crossed species boundaries, in contrast to other hematopoietic growth factors such as [Page 2792] GM-CSF or interleukin-3 (IL-3), which are active on the neutrophilic lineage in a species-specific manner. [...] Nomura et al15 purified native human G-CSF from culture medium conditioned by the tumor cell line CHU-2 and described its properties as a molecule that specifically induced the growth of cells of the neutrophilic granulocyte lineage.15 CLONING OF THE G-CSF GENE Further understanding of the biologic and biochemical properties of G-CSF was greatly facilitated by cloning of the gene encoding G-CSF and the production of recombinant protein for study. 1. Metcalf D: The granulocyte-macrophage colony-stimulating factors. Science 229:16, 1985 2. Metcalf D: The molecular control of cell division, differentiation commitment and maturation in haemopoietic cells. Nature 339:27, 1989 3. Sachs L The molecular control of blood cell development. Science 238:1374, 1987 4. Quesenberry P, Levitt L Hematopoietic stem cells. N Engl J Med 301:755,1979 5. Golde D, Cline M: Regulation of granulopoiesis. N Engl J Med 291:1388,1974 6. Burgess A, Metcalf D: Characterization of a serum factor stimulating the differentiation of myelomonocytic leukemia cells. Int J Cancer 26:647,1980 7. Metcalf D: Clonal extinction of myelomonocytic leukemia cells by serum from mice injected with endotoxin. Int J Cancer 25:225,1980 8. Lotem J, Lipton J, Sachs L Separation of different molecular forms of macrophage and granulocyte-inducing proteins for normal and leukemic myeloid cells. Int J Cancer 25:763,1980 9. Nicola NA, Metcalf D, Matsumoto M, Johnson G R Purification of a factor inducing differentiation in murine myelomonocytic leukemia cells. Identification as granulocyte colony-stimulating factor. J Biol Chem 258:9017,1983 10. Nicola NA, Begley CG, Metcalf D: Identification of the human analogue of a regulator that induces differentiation in murine leukaemic cells. Nature 314:625,1985 14. Souza LM, Boone TC, Gabrilove J, Lai PH, Zsebo KM, Murdock DC, Chazin VR, Bruszewski J, Lu H, Chen KK, Barendt J, Platzer E, Moore MAS, Mertelsmann R, Welte K Recombinant human granulocyte colony-stimulating factor: Effects on normal and leukemic myeloid cells. Science 232:61,1986 15. Nomura H, Imazeki I, Oheda M, Kubota N, Tamura M, Ono M, Ueyama Y, Asano S: Purification and characterization of human granulocyte colony-stimulating actor (G-CSF). EMBO J 5:871,1986 17. Nagata S, Tsuchiya M, Asano S, Kaziro Y, Yamazaki T, Yamamoto 0, Hirata Y, Kubota N, Oheda M, Nomura H, Ono M: Molecular cloning and expression of cDNA for human granulocyte colony-stimulating factor. Nature 319:415,1986 |
Although in this page all but one sentence has been taken - mostly verbatim - from the source Demetri and Griffin (1991) nothing has been marked as a citation, and the source is only given in passing. |
|
| [4.] Arc/Fragment 014 01 - Diskussion Bearbeitet: 26. February 2014, 22:06 Schumann Erstellt: 23. February 2014, 20:39 (Graf Isolan) | Arc, BauernOpfer, Demetri and Griffin 1991, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 14, Zeilen: 1-14, 17-29 |
Quelle: Demetri and Griffin 1991 Seite(n): 2792, Zeilen: 2792: left col. 32-35; right col. 17-25.39-43.48-55; 2793: left col. 13-22 - right col. 1-9 |
|---|---|
| [Further understanding of the biological and biochemical properties of G-CSF was greatly facilitated by cloning of the gene encoding G-CSF and the production of] recombinant protein for study11, 12.
Southern blot analysis of genomic human DNA showed that G-CSF is encoded by a single gene11, 13, and further studies determined that this single gene is located on chromosome 17q11-2214, 15. The genomic structure of the human G-CSF gene was determined by Nagata et al13, revealing that G-CSF gene consists of 5 exons spread over a locus of approximately 2.3 kb (Fig. 1). At the 5’-terminus of the second intron, two donor splice sequences are present in the tandem, only 9 bp apart. The localization of human G-CSF on chromosome 17 differs from that of several other human hematopoietic growth factors such as GM-CSF, IL-3, IL-4 and IL-5, which are clustered on the long arm of the chromosome 516. Following the description of the cDNA for human G-CSF, the murine G-CSF was cloned by crosshybridization with a human G-CSF cDNA probe. The murine G-CSF gene is highly homologous with the human gene, with 69% nucleic acid sequence homology in both coding and non-coding regions, and a 73% sequence homology in the predicted amino acid sequence of the protein17.[...] The human G-CSF gene is distantly related to the IL-6 gene. The number, location and size of the introns and exons that comprise these two genes are similar. Additionally, the amino acid sequences of G-CSF and IL-6 share some localized homology. Between amino acid residues 20 to 85 of G-CSF, the positions of 17 residues match with residues located between positions 28 to 91 of the IL-6 molecule, which yields a sequence homology for this region of 26%3. Additionally, the positions of four cysteine residues are precisely conserved between G-CSF and IL-6 in this region of relative homology. The tertiary structure of G-CSF may be quite similar to that of IL-6, particularly if intra-chain disulfide bridges are similarly located within these molecules. Thus, it is possible that the genes encoding G-CSF and IL-6 may have arisen from a gene duplication event after which they have subsequently diverged. There is no linkage of chromosomal localization between G-CSF and IL-6, because human IL-6 is located at chromosome 7p1519. 3. Demetri GD, Griffin JD. Granulocyte colony-stimulating factor and its receptor. Blood. 1991;78:2791-2808. 11. Nagata S, Tsuchiya M, Asano S, Kaziro Y, Yamazaki T, Yamamoto O, Hirata Y, Kubota N, Oheda M, Nomura H, Ono M. Molecular cloning and expression of cDNA for human granulocyte colony-stimulating factor. Nature. 1986;319:415-418. 12. Souza LM, Boone TC, Gabrilove J, Lai PH, Zsebo KM, Murdock DC, Chazin VR, Bruszewski J, Lu H, Chen KK, et al. Recombinant human granulocyte colony-stimulating factor: effects on normal and leukemic myeloid cells. Science. 1986;232:61-65. 13. Nagata S, Tsuchiya M, Asano S, Yamamoto O, Hirata Y, Kubota N, Oheda M, Nomura H, Yamazaki T. The chromosomal gene structure and two mRNAs for human granulocyte colony-stimulating factor. Embo J. 1986;5:575-581. 14. Le Beau MM, Lemons RS, Carrino JJ, Pettenati MJ, Souza LM, Diaz MO, Rowley JD. Chromosomal localization of the human G-CSF gene to 17q11 proximal to the breakpoint of the t(15;17) in acute promyelocytic leukemia. Leukemia. 1987;1:795-799. 15. Simmers RN, Webber LM, Shannon MF, Garson OM, Wong G, Vadas MA, Sutherland GR. Localization of the G-CSF gene on chromosome 17 proximal to the breakpoint in the t(15;17) in acute promyelocytic leukemia. Blood. 1987;70:330-332. 16. Nicola NA. Hemopoietic cell growth factors and their receptors. Annu Rev Biochem. 1989;58:45-77. 17. Tsuchiya M, Asano S, Kaziro Y, Nagata S. Isolation and characterization of the cDNA for murine granulocyte colony-stimulating factor. Proc Natl Acad Sci U S A. 1986;83:7633-7637. 19. Kishimoto T. The biology of interleukin-6. Blood. 1989;74:1-10. |
[Page 2792]
Further understanding of the biologic and biochemical properties of G-CSF was greatly facilitated by cloning of the gene encoding G-CSF and the production of recombinant protein for study. [...] Southern blot analysis of genomic human DNA showed that human G-CSF is encoded by a single gene,17,18 and further studies determined that this single gene is located on chromosome 17q11-22.20,21 The genomic structure of the human G-CSF gene was determined by Nagata et al.18 The G-CSF gene consists of 5 exons spread over a locus of approximately 2.3 kb. At the 5’-terminus of the second intron, two donor splice sequences are present in tandem, only 9 bp apart. [...] [...] The localization of human G-CSF on chromosome 17 differs from that of several other human hematopoietic growth factors such as GM-CSF, IL-3, IL-4, and IL-5, which are clustered on the long arm of chromosome 5.22 [...] Following the description of the cDNA for human G-CSF, the murine G-CSF gene was cloned by crosshybridization with a human G-CSF cDNA probe under low stringency conditions.23 The murine G-CSF gene is highly homologous with the human gene, with 69% nucleic acid sequence homology in both coding and noncoding regions, and a 73% sequence homology in the predicted amino acid sequence of the protein.23 [Page 2793] The human G-CSF gene is distantly related to the IL-6 gene. The number, location, and size of the introns and exons that comprise these two genes are similar. Additionally, the amino acid sequences of G-CSF and IL-6 share some localized homology. Between amino acid residues 20 to 85 of G-CSF, the positions of 17 residues match with residues located between positions 28 to 91 of the IL-6 molecule, which yields a sequence homology for this region of 26%.26, 27 Additionally, the positions of four cysteine residues are precisely conserved between G-CSF and IL-6 in this region of relative homology. The tertiary structure of G-CSF may be quite similar to that of IL-6, particularly if intrachain disulfide bridges are similarly located within these molecules. Thus, it is possible that the genes encoding G-CSF and IL-6 may have arisen from a gene duplication event from which they have subsequently diverged. There is no linkage of chromosomal localization between G-CSF and IL-6 because the human IL-6 gene is located at chromosome 7p15.28 14. Souza LM, Boone TC, Gabrilove J, Lai PH, Zsebo KM, Murdock DC, Chazin VR, Bruszewski J, Lu H, Chen KK, Barendt J, Platzer E, Moore MAS, Mertelsmann R, Welte K Recombinant human granulocyte colony-stimulating factor: Effects on normal and leukemic myeloid cells. Science 232:61,1986 17. Nagata S, Tsuchiya M, Asano S, Kaziro Y, Yamazaki T, Yamamoto O, Hirata Y, Kubota N, Oheda M, Nomura H, Ono M: Molecular cloning and expression of cDNA for human granulocyte colony-stimulating factor. Nature 319:415,1986 18. Nagata S, Tsuchiya M, Asano S, Yamamoto O, Hirata Y, Kubota N, Oheda M, Nomura H, Yamazaki T: The chromosomal gene structure and two mRNAs for human granulocyte colony-stimulating factor. Embo J 5:575,1986 20. Le Beau M, Lemons R, Carrino J, Pettenati M, Souza L, Diaz M, Rowley J: Chromosomal localization of the human G-CSF gene to 17q11 proximal to the breakpoint of the t(15;17) in acute promyelocytic leukemia. Leukemia 1:795,1987 21. Simmers RN, Webber LM, Shannon MF, Garson OM, Wong G, Vadas MA, Sutherland GR: Localization of the G-CSF gene on chromosome 17 proximal to the breakpoint in the t(15;17) in acute promyelocytic leukemia. Blood 70:330, 1987 22. Nicola N: Hemopoietic cell growth factors and their receptors. Annu Rev Biochem 58:45,1989 23. Tsuchiya M, Asano S, Kaziro Y, Nagata S: Isolation and characterization of the cDNA for murine granulocyte colony-stimulating factor. Proc Natl Acad Sci USA 83:7633,1986 26. Hirano T, Yasukawa K, Harada H, Taga T, Watanabe Y, Matsuda T, Kashiwamura S, Nakajima K, Koyama K, Iwamatsu A, Tsunasawa S, Sakiyama F, Matsui H, Takahara Y, Taniguchi T, Kishimoto T Complementary DNA for a novel human interleukin (BSF-2) that induces B lymphocytes to produce immunoglobulin. Nature 32473,1986 27. Kishimoto T: The biology of interleukin-6. Blood 74:1, 1989 28. Sutherland GR, Baker E, Callen DF, Hyland VJ, Wong G, Clark S, Jones SS, Eglinton LK, Shannon MF, Lopez AF, Vadas MA Interleukin 4 is at 5q31 and interleukin 6 is at 7p15. Hum Genet 79:335,1988 |
Nothing has been marked as a citation. The source is only named once, and then only in passing. |
|
| [5.] Arc/Fragment 015 01 - Diskussion Bearbeitet: 26. February 2014, 01:59 Hindemith Erstellt: 24. February 2014, 22:04 (Hindemith) | Arc, Demetri and Griffin 1991, Fragment, Gesichtet, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 15, Zeilen: 1-1 |
Quelle: Demetri and Griffin 1991 Seite(n): 2793, Zeilen: figure |
|---|---|

Fig. 1 – Molecular organization of the gene encoding human G-CSF, structure of the G-CSF RNA transcript, and secondary structure of the native human G-CSF protein.8 8. Nicola NA, Metcalf D, Matsumoto M, Johnson GR. Purification of a factor inducing differentiation in murine myelomonocytic leukemia cells. Identification as granulocyte colony-stimulating factor. J Biol Chem. 1983;258:9017-9023. |
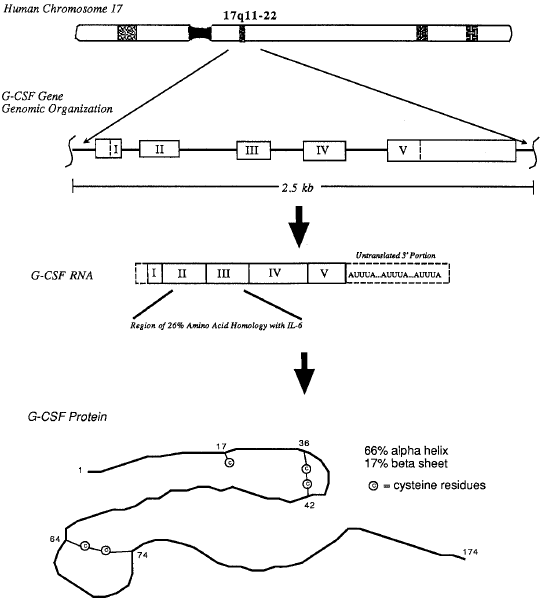
Fig 1. Molecular organization of the gene encoding human G-CSF, structure of the G-CSF RNA transcript, and secondary structure of the native human G-CSF protein. |
The correct source is not given. Neither the image nor the caption can be found in the given source Nicola et al. (1983). |
|
| [6.] Arc/Fragment 016 09 - Diskussion Bearbeitet: 26. February 2014, 23:10 Guckar Erstellt: 25. February 2014, 00:07 (Hindemith) | Arc, Avalos 1996, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 16, Zeilen: 9-16 |
Quelle: Avalos 1996 Seite(n): 761, Zeilen: l.col: 20ff |
|---|---|
| The immature neutrophils that arise from bone marrow follow a process that involves proliferation, differentiation along the granulocyte lineage and terminal maturation into functional neutrophils. Many other hematopoietic growth factors like stem cell factor (SCF), IL-3, GM-CSF, IL-6, have been shown to be positive regulators of granulopoiesis and act at different stages of myeloid cell development. But, G-CSF is unique among the regulators of granulopoiesis, because it not only stimulates the proliferation but also potently induces the terminal maturation of myeloid progenitor cells to neutrophilic granulocytes1.
1. Basu S, Dunn A, Ward A. G-CSF: function and modes of action (Review). Int J Mol Med. 2002;10:3-10. |
Mature neutrophils arise from bone marrow (BM) stem cells following a process involving proliferation, commitment to differentiation along the granulocyte lineage, and terminal maturation.5 Several hematopoietic growth factors including stem cell factor (SCF), interleukin-3 (IL-3), granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-6, and G-CSF have been shown to be positive regulators of granulopoiesis and act at different stages of myeloid cell development.6,7 [...] G-CSF is unique among the regulators of granulopoiesis in that it not only stimulates the proliferation but also potently induces the terminal maturation of myeloid progenitor cells to neutrophilic granulocytes.
[...] |
The source is given before this passage, but without any indication that the text after the reference is taken from it. |
|
| [7.] Arc/Fragment 017 01 - Diskussion Bearbeitet: 27. February 2014, 07:56 Singulus Erstellt: 24. February 2014, 21:49 (Hindemith) | Arc, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Verschleierung, Ward et al 1999 |
|
|
|
| Untersuchte Arbeit: Seite: 17, Zeilen: 1-6 |
Quelle: Ward et al 1999 Seite(n): 14956, Zeilen: r.col: 12-16,19-22 |
|---|---|
| The cytoplasmic region of the G-CSFR can be subdivided into a membrane-proximal domain, which contains two conserved sub-domains known as box 1 and box 2, and a membrane-distal domain, which contains a less conserved box 3 sequence18. In addition, there are four tyrosine residues in the cytoplasmic region of the G-CSFR at positions 704, 729, 744 and 764 of the human receptor, three of which lie in the membrane-distal domain30.
18. Fukunaga R, Ishizaka-Ikeda E, Seto Y, Nagata S. Expression cloning of a receptor for murine granulocyte colony-stimulating factor. Cell Res. 1990;61:341-350. 30. Fukunaga R, Seto Y, Mizushima S, Nagata S. Three Different mRNAs Encoding Human Granulocyte Colony-Stimulating Factor Receptor. Proceedings of the National Academy of Sciences. 1990;87:8702-8706. |
The cytoplasmic region of the G-CSF-R can be subdivided into a membrane-proximal domain, which contains two conserved subdomains known as box 1 and box 2, and a membrane- distal domain, which contains a less-conserved box 3 sequence (5). [...] In addition, there are four tyrosine (Tyr) residues in the cytoplasmic region of the G-CSF-R, at positions 704, 729, 744, and 764 of the human receptor, three of which lie in the membrane-distal domain (28).
5. Fukunaga, R., Ishizaka Ikeda, E., Seto, Y., and Nagata, S. (1990) Cell 61, 341–350 28. Fukunaga, R., Seto, Y., Mizushima, S., and Nagata, S. (1990) Proc. Natl. Acad. Sci. U. S. A. 87, 8702–8706 |
The source is not mentioned anywhere in the paper. Fukunaga et al. (1990) does not contain the documented passage. |
|
| [8.] Arc/Fragment 017 16 - Diskussion Bearbeitet: 26. February 2014, 23:10 Guckar Erstellt: 25. February 2014, 00:21 (Hindemith) | Arc, Avalos 1996, Fragment, Gesichtet, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 17, Zeilen: 16-27 |
Quelle: Avalos 1996 Seite(n): 763, Zeilen: figure |
|---|---|

Fig. 2 – A: Schematic diagram of the structure of the human G-CSFR. The extracellular region (EX) of the G-CSFR contains an Ig-like domain, a cytokine receptor homologous region (CRH) with conserved cysteine (C) residues and the WSXWS motif, and three fibronectin type III (Fn3) domains. The intracellular domain (ID) of the G-CSFR contains three subdomains, designated boxes 1,2, and 3. Numbers correspond to amino acid residues. TM, transmembrane domain; Y, tyrosine residues24. 24. Nagata S, Fukunaga R. Granulocyte colony-stimulating factor and its receptor. Prog Growth Factor Res. 1991;3:131-141. |

Fig 1. Schematic diagram of the structure of the hG-CSFR. The extracellular region (EX) of the G-CSFR contains an Ig-like domain, a cytokine receptor homologous region (CRH) with conserved cysteine (C) residues and the WSXWS motif, and three fibronectin type III (Fn3) domains. The intracellular domain (ID) of the G-CSFR contains three subdomains designated boxes 1,2, and 3 [...]. Numbers correspond to amino acid residues. TM, transmembrane domain; Y, tyrosine residues. |
The source is not given. The copied material cannot be found in Nagata & Fukunaga (1991) |
|
| [9.] Arc/Fragment 017 28 - Diskussion Bearbeitet: 26. February 2014, 22:50 Hindemith Erstellt: 26. February 2014, 00:56 (Hindemith) | Arc, Fragment, Gesichtet, KomplettPlagiat, Nagata Fukunaga 1991, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 17, Zeilen: 28-37 |
Quelle: Nagata Fukunaga 1991 Seite(n): 137, Zeilen: 16ff |
|---|---|
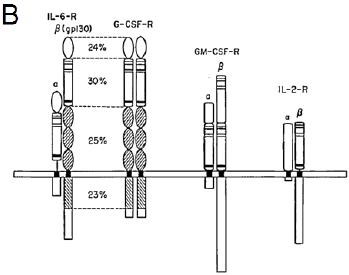
B: Schematic representation of the homo- and hetero-dimeric structures of functional receptors for IL-6, G-CSF, GM-CSF and IL-2. The CRH domains containing the conserved cysteine residues (thin bars) and the WSXWS motif (thick bar) are indicated by shaded boxes. The gp130 (the β-chain of the IL-6 receptor) has a remarkable similarity to the G-CSFR. The numbers indicate the percentage of identical amino acids in each subdomain23. 23. Basu S, Hodgson G, Katz M, Dunn AR. Evaluation of role of G-CSF in the production, survival, and release of neutrophils from bone marrow into circulation. Blood. 2002;100:854-861. |

FIGURE 4. Schematic representation of the homo- and hetero-dimeric structures of functional receptors for IL-6, G-CSF, GM-CSF and IL-2. The CRH domains containing the conserved cysteine residues (thin bars) and the 'WSXWS' motif (thick bar) are indicated by shaded boxes. The gp130 (the β-chain of the IL-6 receptor) has a remarkable similarity to the G-CSF receptor as shown in Fig. 3. The numbers indicate the percentage of identical amino acids in each subdomain. |
The source is not given. None of the copied material can be found in Basu et al. (2002) |
|
| [10.] Arc/Fragment 018 25 - Diskussion Bearbeitet: 26. February 2014, 22:12 Schumann Erstellt: 24. February 2014, 09:37 (Graf Isolan) | Arc, BauernOpfer, Fragment, Gesichtet, SMWFragment, Sampson et al 2007, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 18, Zeilen: 25-31 |
Quelle: Sampson et al 2007 Seite(n): 1 (online source), Zeilen: - |
|---|---|
| G-CSF is an essential growth factor for the optimal production of neutrophils and their precursors. When either G-CSF or its cognate receptor are genetically ablated, the resulting mice are severely neutropenic and susceptible to opportunistic infections20, 40, 41. The loss of G-CSF signalling accounts for the defects on proliferation, differentiation and survival at the progenitor, precursor or terminally differentiated neutrophil stages. The structural functional analysis of the G-CSFR attributes proliferative signalling to the proximal domain (~60 amino acids proximal to the plasma [membrane) and differentiation signalling to the distal domain (~100 amino acids at the C-terminus)29.]
20. Lieschke GJ, Grail D, Hodgson G, Metcalf D, Stanley E, Cheers C, Fowler KJ, Basu S, Zhan YF, Dunn AR. Mice lacking granulocyte colony-stimulating factor have chronic neutropenia, granulocyte and macrophage progenitor cell deficiency, and impaired neutrophil mobilization. Blood. 1994;84:1737-1746. 29. Sampson M, Zhu QS, Corey SJ. Src kinases in G-CSF receptor signaling. Front Biosci. 2007;12:1463-1474. 40. Hermans MH, Ward AC, Antonissen C, Karis A, Lowenberg B, Touw IP. Perturbed granulopoiesis in mice with a targeted mutation in the granulocyte colony-stimulating factor receptor gene associated with severe chronic neutropenia. Blood. 1998;92:32-39. 41. Liu F, Poursine-Laurent J, Link DC. The granulocyte colony-stimulating factor receptor is required for the mobilization of murine hematopoietic progenitors into peripheral blood by cyclophosphamide or interleukin-8 but not flt-3 ligand. Blood. 1997;90:2522-2528. |
2. INTRODUCTION
[...] G-CSF is an essential growth factor for the optimal production of neutrophils and their precursors. When either G-CSF or its cognate receptor is genetically ablated, the resulting mice are severely neutropenic and susceptible to opportunistic infections (3-6). Loss of G-CSF signaling for proliferation, differentiation, and survival at the progenitor, precursor, or terminally differentiated neutrophil stages accounts for these defects. [...]
Structural functional analysis of the G-CSF Receptor attributes proliferative signaling to the proximal domain (~60 amino acids proximal to the plasma membrane) and differentiation to the distal domain (~100 amino acids at the C-terminus) (Figure 1). 3. Hermans, M. H., A. C. Ward, C. Antonissen, A. Karis, B. Lowenberg & I. P. Touw: Perturbed granulopoiesis in mice with a targeted mutation in the granulocyte colony-stimulating factor receptor gene associated with severe chronic neutropenia. Blood, 92, 32-9 (1998)
4. Lieschke, G. J., D. Grail, G. Hodgson, D. Metcalf, E. Stanley, C. Cheers, K. J. Fowler, S. Basu, Y. F. Zhan & A. R. Dunn: Mice lacking granulocyte colony-stimulating factor have chronic neutropenia, granulocyte and macrophage progenitor cell deficiency, and impaired neutrophil mobilization. Blood, 84, 1737-46 (1994)
5. Liu, F., J. Poursine-Laurent & D. C. Link: The granulocyte colony-stimulating factor receptor is required for the mobilization of murine hematopoietic progenitors into peripheral blood by cyclophosphamide or interleukin-8 but not flt-3 ligand. Blood, 90, 2522-8 (1997)
6. Liu, F., H. Y. Wu, R. Wesselschmidt, T. Kornaga & D. C. Link: Impaired production and increased apoptosis of neutrophils in granulocyte colony-stimulating factor receptor-deficient mice. Immunity, 5, 491-501 (1996) 10.1016/S1074-7613 (00)80504-X |
Though the source is named in the end nothing has been marked as a citation. Also the references to the literature have been taken from the source. |
|
| [11.] Arc/Fragment 019 01 - Diskussion Bearbeitet: 26. February 2014, 10:44 Hindemith Erstellt: 24. February 2014, 21:39 (Graf Isolan) | Arc, BauernOpfer, Fragment, Gesichtet, SMWFragment, Sampson et al 2007, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 19, Zeilen: 1-34 |
Quelle: Sampson et al 2007 Seite(n): 1 (online source), Zeilen: - |
|---|---|
| [The structural functional analysis of the G-CSFR attributes proliferative signalling to the proximal domain (~60 amino acids proximal to the plasma] membrane) and differentiation signalling to the distal domain (~100 amino acids at the C-terminus)29.
Current models of cytokine receptor signalling, including that for G-CSFR, assign a critical signal transduction role to Janus kinases (Jak). Although Jak2-deficient mice display major defects in IL-3 and GM-CSF signalling, mice deficient in either Jak1 or Jak2 have intact G-CSFR signalling42, 43. The Src family of proto-oncogenic tyrosine kinases seems to have a more important role in transducing G-CSF-induced cell cycle progression. Since Src kinases have a wider range of physiological substrates than do the Jaks, which primarily affect the STAT proteins, Src kinases may play a more important role in G-CSF-mediated cell survival and metabolism. These pathways that involve Erk1/2 or PI3-kinase may contribute to G-CSF-induced proliferation, differentiation, survival and cytoskeletal reorganization29. Multiple protein tyrosine kinases (PTK) (e.g., Jak2 and Src) probably phosphorylate the tyrosine residues (tyr704, tyr729, tyr744 and tyr764) of the G-CSFR44. When phosphorylated, the phosphotyrosine residues serve as docking sites for signalling proteins containing phosphotyrosine binding domains (e.g., SH2 or PTB). Recruitment of these signalling proteins serves to diversify and inactivate G-CSFR signal. Diversification involves recruitment of the STAT transcription factors and Ras/Erk1/2 and PI3-kinase pathways (Fig. 3). Tyr704 can be phosphorylated by the Jaks and then serve as a docking site for the SH2 domains of STAT proteins45. Resembling that site, when phosphorylated, tyr729 may also serve as a docking site for the SH2 domain of STAT. Tyr764 favours the Src kinase and the Src SH2 domain46. This site is also the preferred binding site for the SH2 domains of Grb2 and is functionally coupled to Shc and the SH2-containing tyrosine phosphatise-2 (Shp-2)47. Grb2 also interacts with Gab2, which leads to PI3-kinase activity48. Tyr764 is also functionally coupled to Ras activation and Jun kinase49. Thus, phosphor-tyr764 can transduce several different signals, with both positive and negative effects on growth. Substrate availability and sustained activation may determine functional outcome. In their phosphorylated states, tyr 744 and tyr 729 may serve as docking sites for cytokine inducible SH2 protein/suppressor of cytokine signalling (SOCS) and SH2-containing inositol phosphatese (SHIP)50. Both molecules are negative regulators of Jak-STAT and PI3-kinase, respectively. The C-terminal domain also recruits SH2-containing tyrosine phosphatise-1 (Shp-1), which dephosphorylates positive signalling molecules such as Lyn and STAT51. According to what was said, while there may be a crosstalk between Src and Jak signalling pathways, [each kinase can trigger a stereotyped response, e.g., Jak-STAT and Src-Ras/Pi3-kinase29.] 29. Sampson M, Zhu QS, Corey SJ. Src kinases in G-CSF receptor signaling. Front Biosci. 2007;12:1463-1474. 42. Parganas E, Wang D, Stravopodis D, Topham DJ, Marine JC, Teglund S, Vanin EF, Bodner S, Colamonici OR, van Deursen JM, Grosveld G, Ihle JN. Jak2 is essential for signaling through a variety of cytokine receptors. Cell. 1998;93:385-395. 43. Rodig SJ, Meraz MA, White JM, Lampe PA, Riley JK, Arthur CD, King KL, Sheehan KC, Yin L, Pennica D, Johnson EM, Jr., Schreiber RD. Disruption of the Jak1 gene demonstrates obligatory and nonredundant roles of the Jaks in cytokine-induced biologic responses. Cell. 1998;93:373-383. 44. Corey SJ, Dombrosky-Ferlan PM, Zuo S, Krohn E, Donnenberg AD, Zorich P, Romero G, Takata M, Kurosaki T. Requirement of Src kinase Lyn for induction of DNA synthesis by granulocyte colony-stimulating factor. J Biol Chem. 1998;273:3230-3235. 45. de Koning JP, Dong F, Smith L, Schelen AM, Barge RM, van der Plas DC, Hoefsloot LH, Lowenberg B, Touw IP. The membrane-distal cytoplasmic region of human granulocyte colony-stimulating factor receptor is required for STAT3 but not STAT1 homodimer formation. Blood. 1996;87:1335-1342. 46. Songyang Z, Shoelson SE, Chaudhuri M, Gish G, Pawson T, Haser WG, King F, Roberts T, Ratnofsky S, Lechleider RJ, et al. SH2 domains recognize specific phosphopeptide sequences. Cell. 1993;72:767-778. 47. de Koning JP, Schelen AM, Dong F, van Buitenen C, Burgering BM, Bos JL, Lowenberg B, Touw IP. Specific involvement of tyrosine 764 of human granulocyte colony-stimulating factor receptor in signal transduction mediated by p145/Shc/GRB2 or p90/GRB2 complexes. Blood. 1996;87:132-140. 48. Zhu QS, Robinson LJ, Roginskaya V, Corey SJ. G-CSF-induced tyrosine phosphorylation of Gab2 is Lyn kinase dependent and associated with enhanced Akt and differentiative, not proliferative, responses. Blood. 2004;103:3305-3312. 49. Rausch O, Marshall CJ. Tyrosine 763 of the murine granulocyte colony-stimulating factor receptor mediates Ras-dependent activation of the JNK/SAPK mitogen-activated protein kinase pathway. Mol Cell Biol. 1997;17:1170-1179. 50. Hunter MG, Jacob A, O'Donnell L C, Agler A, Druhan LJ, Coggeshall KM, Avalos BR. Loss of SHIP and CIS recruitment to the granulocyte colony-stimulating factor receptor contribute to hyperproliferative responses in severe congenital neutropenia/acute myelogenous leukemia. J Immunol. 2004;173:5036-5045. |
3. THE G-CSF RECEPTOR
[...] Current models of cytokine receptor signaling, including that for the G-CSF Receptor, assign the critical signal transduction role to the Janus kinases. [...] In distinction to Jak2-deficient mice that display major defects in IL-3, GM-CSF, Epo, and TPO signaling, mice deficient in either Jak1 or Jak2 have intact G-CSF Receptor signaling (22-24). As discussed below, Src kinases have a non-redundant function in transducing G-CSF-induced cell cycle progression. Src kinases have a wider range of physiological substrates than do the Jaks, which primarily affect the STAT proteins (Figure 1). Src kinases may also play a role in G-CSF-mediated survival and metabolism. These pathways that involve the Erk1/2 or PI 3-kinase may contribute to G-CSF-induced differential and cytoskeletal reorganization. [...] 6. TYROSINE PHOSPHORYLATION OF THE G-CSF RECEPTOR Structural functional analysis of the G-CSF Receptor attributes proliferative signaling to the proximal domain (~60 amino acids proximal to the plasma membrane) and differentiation to the distal domain (~100 amino acids at the C-terminus) (Figure 1). [...] Multiple PTK (e.g. Jak2 and Src) probably phosphorylate the tyrosine residues (Tyr704, Tyr729, Tyr744, and Tyr764) of the G-CSF Receptor (16). [...] When phosphorylated, the phosphotyrosine residues serve as docking sites for signaling proteins containing a phosphotyrosine binding domains (e.g. SH2 or PTB). Recruitment of these signaling proteins serves to diversify and inactivate G-CSF Receptor's signal. Diversification involves recruitment of the STAT transcription factors and Ras/Erk1/2 and PI 3'kinase pathways (Figure 3). Y704VLQ fits the YXXQ motif, which can be phosphorylated by the Jaks and then serve as a docking site for the SH2 domains of STAT proteins (17). Resembling that site, when phosphorylated, Y729GQL may also serve as a docking site for the SH2 domain of STAT. Y764ENL best approximates the YEEI/L motif favored by both the Src kinase and the Src SH2 domain (56, 60). This site is also the preferred binding site (i.e. YpEN) for the SH2 domain of Grb2 and is functionally coupled to Shc and the SH2-containing tyrosine phosphatase-2 (Shp-2)(18). Grb2 also interacts with Gab2, which leads to PI 3'-kinase activity (61). Tyr764 is also functionally coupled to Ras activation and Jun kinase (62). Thus, phospho-Tyr764 can transduce several different signals with both positive and negative effects on growth. Substrate availability and sustained activation may determine functional outcome. In their phosphorylated states, Y744LRC and Y729GQL may serve as docking sites for cytokine inducible SH2 protein (CIS)/suppressor of cytokine signaling (SOCS) and SH2-containing inositol phosphatase (SHIP) (63-66). Both molecules are negative regulators of Jak-STAT and PI 3'-kinase, respectively. The C-terminal domain also recruits SH2-containing tyrosine phosphatase-1 (Shp-1), which desphosphorylates positive signaling molecules such as Lyn and STAT (67). 7. INTRACELLULAR SIGNALING IN MYELOID CELLS [...] [...] While there may be cross-talk between Src and Jak signaling pathways, each kinase can trigger a stereotyped response, e.g. Jak-STAT and Src-Ras/PI 3-kinase. 16. Corey, S. J., P. M. Dombrosky-Ferlan, S. Zuo, E. Krohn, A. D. Donnenberg, P. Zorich, G. Romero, M. Takata & T. Kurosaki: Requirement of Src kinase Lyn for induction of DNA synthesis by granulocyte colony-stimulating factor. J Biol Chem, 273, 3230-5 (1998) 10.1074/jbc.273.6.3230 17. de Koning, J. P., F. Dong, L. Smith, A. M. Schelen, R. M. Barge, D. C. van der Plas, L. H. Hoefsloot, B. Lowenberg & I. P. Touw: The membrane-distal cytoplasmic region of human granulocyte colony-stimulating factor receptor is required for STAT3 but not STAT1 homodimer formation. Blood, 87, 1335-42 (1996) 18. de Koning, J. P., A. M. Schelen, F. Dong, C. van Buitenen, B. M. Burgering, J. L. Bos, B. Lowenberg & I. P. Touw: Specific involvement of tyrosine 764 of human granulocyte colony-stimulating factor receptor in signal transduction mediated by p145/Shc/GRB2 or p90/GRB2 complexes. Blood, 87, 132-40 (1996) 22. Parganas, E., D. Wang, D. Stravopodis, D. J. Topham, J. C. Marine, S. Teglund, E. F. Vanin, S. Bodner, O. R. Colamonici, J. M. van Deursen, G. Grosveld & J. N. Ihle: Jak2 is essential for signaling through a variety of cytokine receptors. Cell, 93, 385-95 (1998) 10.1016/S0092-8674 (00)81167-8 23. Rodig, S. J., M. A. Meraz, J. M. White, P. A. Lampe, J. K. Riley, C. D. Arthur, K. L. King, K. C. Sheehan, L. Yin, D. Pennica, E. M. Johnson, Jr. & R. D. Schreiber: Disruption of the Jak1 gene demonstrates obligatory and nonredundant roles of the Jaks in cytokine-induced biologic responses. Cell, 93, 373-83 (1998) 10.1016/S0092-8674 (00)81166-6 24. Neubauer, H., A. Cumano, M. Muller, H. Wu, U. Huffstadt & K. Pfeffer: Jak2 deficiency defines an essential developmental checkpoint in definitive hematopoiesis. Cell, 93, 397-409 (1998) 10.1016/S0092-8674 (00)81168-X 56. Songyang, Z., S. E. Shoelson, M. Chaudhuri, G. Gish, T. Pawson, W. G. Haser, F. King, T. Roberts, S. Ratnofsky, R. J. Lechleider & et al.: SH2 domains recognize specific phosphopeptide sequences. Cell, 72, 767-78 (1993) 61. Zhu, Q. S., L. J. Robinson, V. Roginskaya & S. J. Corey: G-CSF-induced tyrosine phosphorylation of Gab2 is Lyn kinase dependent and associated with enhanced Akt and differentiative, not proliferative, responses. Blood, 103, 3305-12 (2004) 62. Rausch, O. & C. J. Marshall: Tyrosine 763 of the murine granulocyte colony-stimulating factor receptor mediates Ras-dependent activation of the JNK/SAPK mitogen-activated protein kinase pathway. Mol Cell Biol, 17, 1170-9 (1997) 63. Hunter, M. G., A. Jacob, C. O'Donnell L, A. Agler, L. J. Druhan, K. M. Coggeshall & B. R. Avalos: Loss of SHIP and CIS recruitment to the granulocyte colony-stimulating factor receptor contribute to hyperproliferative responses in severe congenital neutropenia/acute myelogenous leukemia. J Immunol, 173, 5036-45 (2004) 64. van de Geijn, G. J., J. Gits, L. H. Aarts, C. Heijmans-Antonissen & I. P. Touw: G-CSF receptor truncations found in SCN/AML relieve SOCS3-controlled inhibition of STAT5 but leave suppression of STAT3 intact. Blood, 104, 667-74 (2004) 65. Hermans, M. H., G. J. van de Geijn, C. Antonissen, J. Gits, D. van Leeuwen, A. C. Ward & I. P. Touw: Signaling mechanisms coupled to tyrosines in the granulocyte colony-stimulating factor receptor orchestrate G-CSF-induced expansion of myeloid progenitor cells. Blood, 101, 2584-90 (2003) 66. van de Geijn, G. J., J. Gits & I. P. Touw: Distinct activities of suppressor of cytokine signaling (SOCS) proteins and involvement of the SOCS box in controlling G-CSF signaling. J Leukoc Biol, 76, 237-44 (2004) |
Though the source is named in passing nothing has been marked as a citation. Note that also all the references to the literature have been taken from the source. |
|
| [12.] Arc/Fragment 020 03 - Diskussion Bearbeitet: 26. February 2014, 23:55 Hindemith Erstellt: 24. February 2014, 22:23 (Hindemith) | Arc, Fragment, Gesichtet, SMWFragment, Sampson et al 2007, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 20, Zeilen: 3-4 |
Quelle: Sampson et al 2007 Seite(n): 1 (online source), Zeilen: figures section |
|---|---|
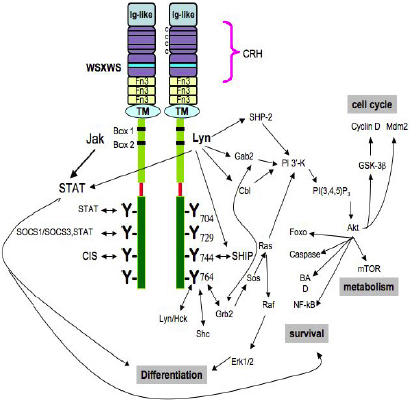
Fig. 3 – Diverse intracellular signalling pathways of the G-CSF Receptor. The cytoplasmic domain of the receptor contains Box 1 and Box 2 and Tyr704 in the proximal domain, necessary for mitogenesis. When phosphorylated, the four tyrosine residues serve as docking sites for SH2-containing proteins. Neither the structural basis of how cytosolic PTKs, such as Jak and Lyn, associate with the receptor, nor the specificity or redundancy of each tyrosine residue’s kinase are known28. 28. Basu S, Hodgson G, Zhang H-H, Katz M, Quilici C, Dunn AR. "Emergency" granulopoiesis in G-CSF-deficient mice in response to Candida albicans infection. Blood. 2000;95:3725-3733. |
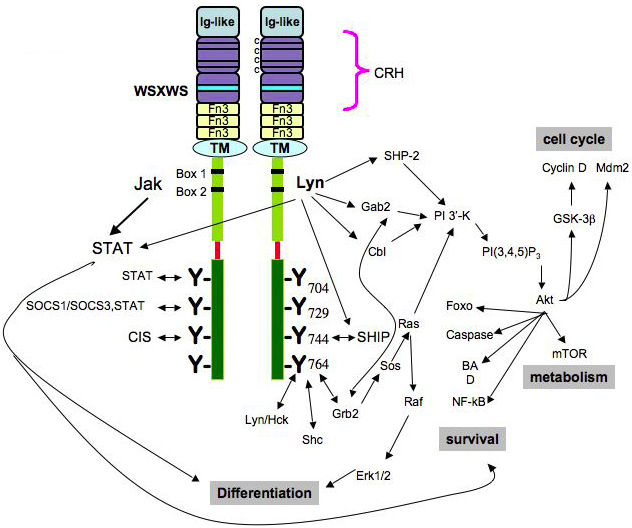
Figure 1. Intracellular signaling pathways of the G-CSF Receptor. [...] The cytoplasmic domain of the receptor contains Box 1 and Box 2 and Tyr704 in the proximal domain, which is necessary for mitogenesis. [...] When phosphorylated, the four tyrosine residues serve as docking sites for SH2-containing proteins. [...] Neither the structural basis of how cytosolic PTKs, such as Jak and Lyn, associate with the receptor, nor the the specificity or redundancy of each tyrosine residue's kinase are known. |
The source is not mentioned here. Neither image nor caption can be found in Basu et al. (2000). |
|
| [13.] Arc/Fragment 021 11 - Diskussion Bearbeitet: 26. February 2014, 20:44 Graf Isolan Erstellt: 24. February 2014, 18:44 (Hindemith) | Arc, Fragment, Gesichtet, Quinn and Gauss 2004, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 21, Zeilen: 11-21 |
Quelle: Quinn and Gauss 2004 Seite(n): 760, Zeilen: r.col: 16ff |
|---|---|
| Neutrophil granulocytes (a.k.a., polymorphonuclear leukocytes) are normally found circulating in the bloodstream (half-life of ≅7 h) and migrating through tissues (2–3 days), devoting their short lifetime to surveillance55. During an infection, the neutrophil lifespan is increased, and large numbers of neutrophils are rapidly recruited to the site(s) of infection where they function to destroy invading pathogens. In this manner, neutrophils serve as one of the body’s first lines of defence against infection. These cells use an extraordinary array of oxygen-dependent and oxygen-independent microbicidal weapons to destroy and remove infectious agents56. Oxygen-dependent mechanisms involve the production of ROS, which can be microbicidal57, and oxygen-independent mechanisms include most other neutrophil functions, such as chemotaxis, phagocytosis, degranulation, and release of lytic enzymes and bactericidal peptides56.
55. Steinfeld JL. Principles of Hematology. Cancer Res. 1967;27:208-a-. 56. Witko-Sarsat V, Rieu P, Descamps-Latscha B, Lesavre P, Halbwachs-Mecarelli L. Neutrophils: Molecules, Functions and Pathophysiological Aspects. Lab Invest. 2000;80:617-653. 57. Roos D, Bruggena Rv, Meischl C. Oxidative killing of microbes by neutrophils. Microbes and Infection. 2003;5:1307-1315. |
Neutrophils (a.k.a., polymorphonuclear leukocytes) are normally found circulating in the bloodstream (circulating half-life of ~7 h) and migrating through tissues (2–3 days) and devote their short lifetime to surveillance [3]. However, during an infection, the neutrophil lifespan is increased, and large numbers of neutrophils are rapidly recruited to the site(s) of infection where they function to destroy invading pathogens. In this capacity, neutrophils serve as one of the body’s first lines of defense against infection. These cells use an extraordinary array of oxygen-dependent and oxygen-independent microbicidal weapons to destroy and remove infectious agents [4]. Oxygen-dependent mechanisms involve the production of reactive oxygen species (ROS), which can be microbicidal [5], and oxygen-independent mechanisms include most other neutrophil functions, such as chemotaxis, phagocytosis, degranulation, and release of lytic enzymes and bactericidal peptides (reviewed in ref. [4]).
3. Haen, P. J. (1995) Principles of Hematology, Dubuque, IA, Wm. C. Brown. 4. Witko-Sarsat, V., Rieu, P., Descamps-Latscha, B., Lesavre, P., Halbwachs-Mecarelli, L. (2000) Neutrophils: molecules, functions and pathophysiological aspects. Lab. Invest. 80, 617–653. 5. Roos, D., Van Bruggen, R., Meischl, C. (2003) Oxidative killing of microbes by neutrophils. Microbes Infect. 5, 1307–1315. |
The source is not given here. |
|
| [14.] Arc/Fragment 021 22 - Diskussion Bearbeitet: 26. February 2014, 22:17 Schumann Erstellt: 23. February 2014, 06:20 (Hindemith) | Arc, BauernOpfer, Bedard Krause 2007, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 21, Zeilen: 22-30 |
Quelle: Bedard Krause 2007 Seite(n): 246, Zeilen: l.col: 9ff |
|---|---|
| ROS are oxygen-derived small molecules, including oxygen radicals [superoxide (O2•–), hydroxyl (•OH), peroxyl (RO2•), and alkoxyl (RO•) radicals] and certain non-radicals that are either oxidizing agents and/or are easily converted into radicals, such as hypochlorous acid (HOCl), ozone (O3), singlet oxygen (1O2), and hydrogen peroxide (H2O2). ROS generation is generally a cascade of reactions that starts with the production of superoxide. Superoxide rapidly dismutates to hydrogen peroxide either spontaneously (at low pH) or catalyzed by superoxide dismutase (SOD). Other elements in the cascade of ROS generation include the reaction of superoxide with nitric oxide to form peroxynitrite (ONOO–), the peroxidase-catalyzed formation of hypochlorous acid [(HOCl) from hydrogen peroxide, and the iron-catalyzed Fenton reaction leading to the generation of hydroxyl radical58.]
Thannickal VJ, Fanburg BL. Reactive oxygen species in cell signaling. Am J Physiol Lung Cell Mol Physiol. 2000;279:L1005-1028. |
Reactive oxygen species (ROS) are oxygen-derived small molecules, including oxygen radicals [superoxide (O2•–), hydroxyl (•OH), peroxyl (RO2•), and alkoxyl (RO•)] and certain nonradicals that are either oxidizing agents and/or are easily converted into radicals, such as hypochlorous acid (HOCl), ozone (O3), singlet oxygen (1O2), and hydrogen peroxide (H2O2). Nitrogen-containing oxidants, such as nitric oxide, are called reactive nitrogen species (RNS). ROS generation is generally a cascade of reactions that starts with the production of superoxide. Superoxide rapidly dismutates to hydrogen peroxide either spontaneously, particularly at low pH or catalyzed by superoxide dismutase. Other elements in the cascade of ROS generation include the reaction of superoxide with nitric oxide to form peroxynitrite, the peroxidase-catalyzed formation of hypochlorous acid from hydrogen peroxide, and the iron-catalyzed Fenton reaction leading to the generation of hydroxyl radical (468, 874).
468. Klebanoff SJ. Oxygen metabolism and the toxic properties of phagocytes. Ann Intern Med 93: 480–489, 1980. 874. Thannickal VJ, Fanburg BL. Reactive oxygen species in cell signaling. Am J Physiol Lung Cell Mol Physiol 279: L1005–L1028, 2000. |
The source is mentioned further down on the next page, without any indication that everything including references to the literature are taken from it. To be continued on the next page: Arc/Fragment_022_01 |
|
| [15.] Arc/Fragment 022 01 - Diskussion Bearbeitet: 26. February 2014, 22:19 Schumann Erstellt: 23. February 2014, 06:35 (Hindemith) | Arc, BauernOpfer, Bedard Krause 2007, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 22, Zeilen: 1-13 |
Quelle: Bedard Krause 2007 Seite(n): 246, Zeilen: l.col: 21ff |
|---|---|
| [Other elements in the cascade of ROS generation include the reaction of superoxide with nitric oxide to form peroxynitrite (ONOO-), the peroxidase-catalyzed formation of hypochlorous acid] (HOCl) from hydrogen peroxide, and the iron-catalyzed Fenton reaction leading to the generation of hydroxyl radical58. ROS avidly interact with a large number of molecules including other small inorganic molecules as well as proteins, lipids, carbohydrates, and nucleic acids. Through such interactions, ROS may irreversibly destroy or alter the function of the target molecule and consequently, ROS have been increasingly identified as major contributors to damage in biological organisms. However, ROS are involved not only in cellular damage and killing of pathogens, but also in a large number of reversible regulatory signalling processes in virtually all cells and tissues59. The physiological generation of ROS can occur as a result of other biological reactions. For example, ROS generation occurs as a byproduct in the mitochondria, peroxisomes, cytochrome P-450, and other cellular elements58. The phagocyte NADPH oxidase was the first identified example of a system that generates ROS not as a byproduct, but rather as the primary function of this enzyme system59.
58. Thannickal VJ, Fanburg BL. Reactive oxygen species in cell signaling. Am J Physiol Lung Cell Mol Physiol. 2000;279:L1005-1028. 59. Bedard K, Krause K-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007;87:245-313. |
Other elements in the cascade of ROS generation include the reaction of superoxide with nitric oxide to form peroxynitrite, the peroxidase-catalyzed formation of hypochlorous acid from hydrogen peroxide, and the iron-catalyzed Fenton reaction leading to the generation of hydroxyl radical (468, 874).
ROS avidly interact with a large number of molecules including other small inorganic molecules as well as proteins, lipids, carbohydrates, and nucleic acids. Through such interactions, ROS may irreversibly destroy or alter the function of the target molecule. Consequently, ROS have been increasingly identified as major contributors to damage in biological organisms. [...] In fact, ROS are involved not only in cellular damage and killing of pathogens, but also in a large number of reversible regulatory processes in virtually all cells and tissues. [...] [...] The physiological generation of ROS can occur as a byproduct of other biological reactions. ROS generation as a byproduct occurs with mitochondria, peroxisomes, cytochrome P-450, and other cellular elements (50, 307, 314, 356, 588, 636, 715, 791, 874). However, the phagocyte NADPH oxidase was the first identified example of a system that generates ROS not as a byproduct, but rather as the primary function of the enzyme system. [...] 874. Thannickal VJ, Fanburg BL. Reactive oxygen species in cell signaling. Am J Physiol Lung Cell Mol Physiol 279: L1005–L1028, 2000. [...] |
The copied text starts on the previous page: see: Arc/Fragment_021_22. The source is given twice, but still it is not clear to the reader that everything is taken more or less verbatim from the source, including the reference to Thannickal & Fanburg (2000). |
|
| [16.] Arc/Fragment 022 15 - Diskussion Bearbeitet: 26. February 2014, 21:34 Graf Isolan Erstellt: 24. February 2014, 18:56 (Hindemith) | Arc, BauernOpfer, Fragment, Gesichtet, Quinn and Gauss 2004, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 22, Zeilen: 15-24 |
Quelle: Quinn and Gauss 2004 Seite(n): 761, Zeilen: l.col: 23ff |
|---|---|
| In general, the best characterized plasma membrane oxidase is the phagocytic NADPH oxidase, a multicomponent enzyme composed of four oxidase-specific proteins (p22phox, p47phox, p67phox, and gp91phox) and a GTPase (Rac1/2). One other oxidase-specific protein (p40phox) and a second GTPase (Rap1A) have also been shown to play roles in regulating oxidase activity; however, their specific functions are still not well understood60. Originally, the nomenclature for the various components differed throughout the literature (Table 1); however, the generally accepted nomenclature for the phagocyte oxidase specific components now includes the suffix phox, which refers to phagocyte oxidase61. The only one exception is gp91phox, which has also been named NADPH oxidase 2 (Nox2)62.
60. Quinn MT, Gauss KA. Structure and regulation of the neutrophil respiratory burst oxidase: comparison with nonphagocyte oxidases. J Leukoc Biol. 2004;76:760-781. 61. Babior BM. NADPH oxidase. Current Opinion in Immunology. 2004;16:42-47. 62. Lambeth JD, Cheng G, Arnold RS, Edens WA. Novel homologs of gp91phox. Trends Biochem Sci. 2000;25:459-461. |
It is now generally accepted that the core NADPH oxidase enzyme is composed of four oxidase-specific proteins (p22phox, p47phox, p67phox, and gp91phox) and a GTPase (Rac1/2). One other oxidase-specific protein (p40phox) and a second GTPase (Rap1A) have also been shown to play roles in regulating oxidase activity; however, their specific functions are still not well understood. Originally, the nomenclature for the various components differed throughout the literature; however, the generally accepted nomenclature for the phagocyte oxidasespecific components now includes the suffix phox, which refers to phagocyte oxidase [10]. The one exception is gp91phox, which has also been named NADPH oxidase 2 (Nox2) [11].
10. Babior, B. M. (1991) The respiratory burst oxidase and the molecular basis of chronic granulomatous disease. Am. J. Hematol. 37, 263–266. 11. Lambeth, J. D., Cheng, G., Arnold, R. S., Edens, W. A. (2000) Novel homologs of gp91phox. Trends Biochem. Sci. 25, 459–461. |
The source is mentioned somewhere in the middle, but it is not clear to the reader that the entire passage is taken from it, including a reference to the literature. |
|
| [17.] Arc/Fragment 023 01 - Diskussion Bearbeitet: 26. February 2014, 22:46 WiseWoman Erstellt: 23. February 2014, 07:16 (Hindemith) | Arc, Bedard Krause 2007, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 23, Zeilen: 1-5 |
Quelle: Bedard Krause 2007 Seite(n): 248, 252, Zeilen: 248: 1ff; 252: 1ff |
|---|---|
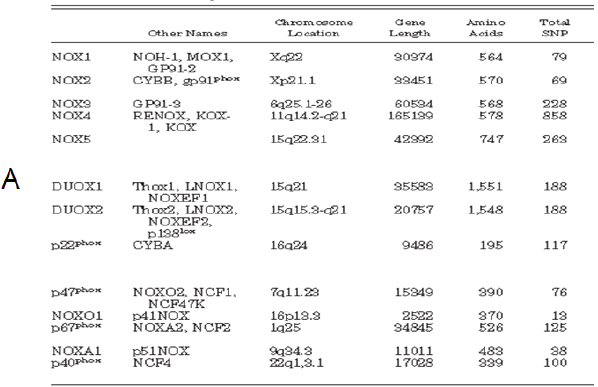
Table 1 – A: Nox enzymes and subunits: alternative names, chromosomal localization, gene length, number of amino acids, and total number of single-nucleotide polymorphisms (total SNP). B: Tissue distribution of Nox enzymes: Nox enzymes are expressed in a small number of tissues at high levels (readily detected by Northern blotting) but show intermediate- to low-level expression in many other tissues57. 57. Roos D, Bruggena Rv, Meischl C. Oxidative killing of microbes by neutrophils. Microbes and Infection. 2003;5:1307-1315. |
[page 248]
Alternative names, chromosomal localization, gene length, number of amino acids, total number of single-nucleotide polymorphisms (total SNP), number of coding nonsynonymous single-nucleotide polymorphisms (cnSNP), amino acid changes in cnSNPs, and nucleotide substitutions in cnSNPs are shown. The number of SNPs is based on presently available NCBI databank entries. The position of the nucleotide substitution is given relative to the start codon (207). * C214T in p22phox was originally called C242T (411), a name still widely used in the literature. [page 252]
NOX enzymes are expressed in a small number of tissues at high levels (readily detected by Northern blotting) but show intermediate- to low-level expression in many other tissues. 207. Den Dunnen JT, Antonarakis SE. Nomenclature for the description of human sequence variations. Hum Genet 109: 121–1 411. Inoue N, Kawashima S, Kanazawa K, Yamada S, Akita H, Yokoyama M. Polymorphism of the NADH/NADPH oxidase p22phox gene in patients with coronary artery disease. Circulation 97: 135–137, 1998. |
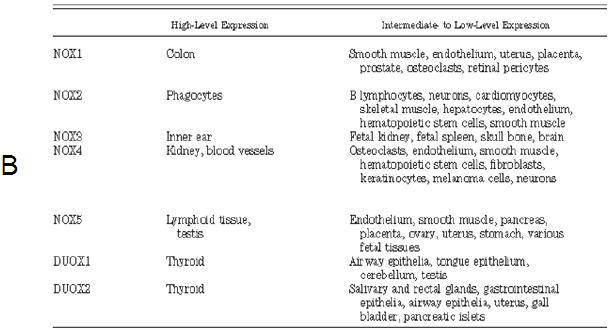
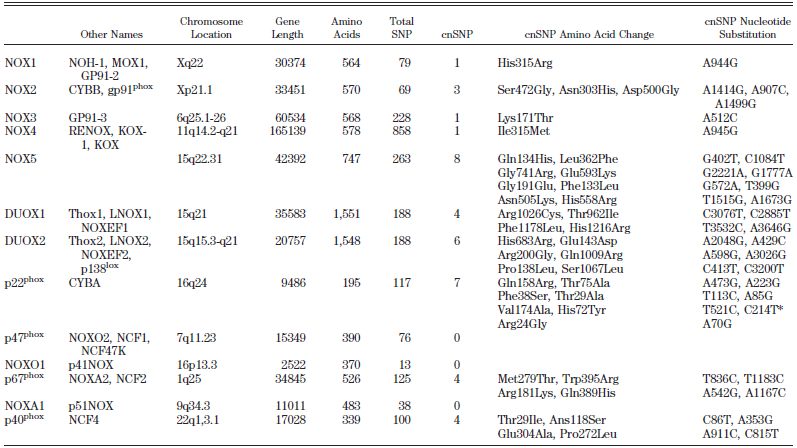
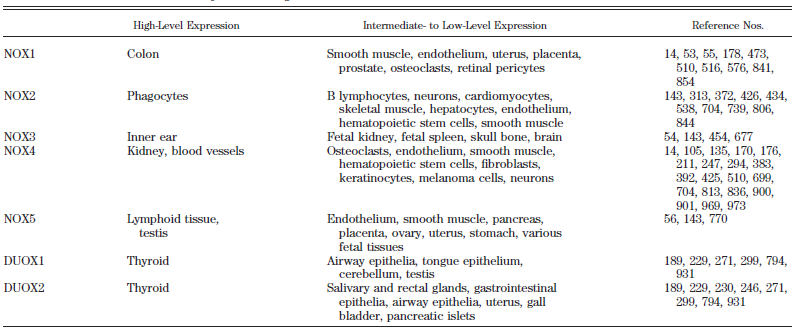
The two tables are copied verbatim from the source, which is not referenced. The source given, Roos et al. (2003). does not contain any of the copied material. |
|
| [18.] Arc/Fragment 024 06 - Diskussion Bearbeitet: 26. February 2014, 23:16 Schumann Erstellt: 23. February 2014, 07:36 (Hindemith) | Arc, BauernOpfer, Bedard Krause 2007, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 24, Zeilen: 6-19 |
Quelle: Bedard Krause 2007 Seite(n): 249, 250, Zeilen: 249: l.col: 16ff |
|---|---|
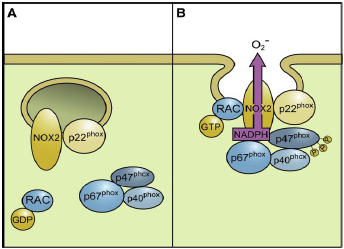
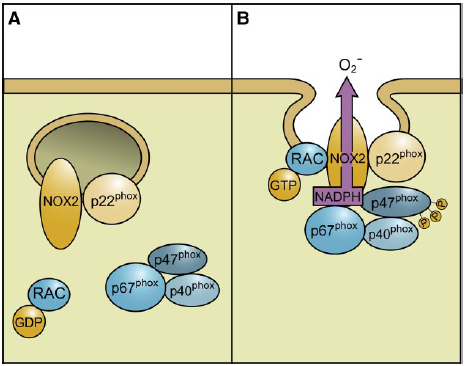
| The activation of Nox2 occurs through a complex series of protein/protein interactions, where Nox2 constitutively associates with p22phox, forming a heterodimeric complex known as cytochrome b558 (Cyt b558)58. NADPH oxidase activation requires translocation of cytosolic factors to the Nox2/p22phox complex and the present working model functions in the following way (Fig. 4): First, phosphorylation of p47phox leads to a conformational change allowing its interaction with p22phox. It is thought that p47phox organizes the translocation of other cytosolic factors, hence its designation as “organizer subunit.” The relocalization of p47phox to the membrane brings the “activator subunit” p67phox into contact with Nox2 and also brings the small subunit p40phox to the complex. Finally, the GTPase Rac interacts with Nox2 via a two-step mechanism involving an initial direct interaction with Nox2, followed by a subsequent interaction with p67phox. Once assembled, the complex is active and generates superoxide by transferring an electron from NADPH in the cytosol to oxygen on the luminal or extracellular space59.
Fig. 4 - Assembly of the phagocyte NADPH oxidase Nox2. Nox2 and p22phox are found primarily in the membrane of intracellular vesicles. Upon activation, there is an exchange of GDP for GTP on Rac leading to its activation. Phosphorylation of the cytosolic p47phox subunit leads to conformational changes allowing interaction with p22phox. The movement of p47phox brings with it the other cytoplasmic subunits, p67phox and p40phox, to form the active Nox2 enzyme complex. Once activated, there is a fusion of Nox2-containing vesicles with the plasma membrane or the phagosomal membrane. The active enzyme complex transports electrons from cytoplasmic NADPH to extracellular or phagosomal oxygen to generate superoxide57 57. Roos D, Bruggena Rv, Meischl C. Oxidative killing of microbes by neutrophils. Microbes and Infection. 2003;5:1307-1315. 58. Thannickal VJ, Fanburg BL. Reactive oxygen species in cell signaling. Am J Physiol Lung Cell Mol Physiol. 2000;279:L1005-1028. 59. Bedard K, Krause K-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007;87:245-313. |
The activation of NOX2 occurs through a complex series of protein/protein interactions (Fig. 2; for more detailed recent reviews, see Refs. 328, 652, 844). NOX2 constitutively associates with p22phox. [...] Activation of NOX2 requires translocation of cytosolic factors to the NOX2/p22phox complex (Fig. 3). The present working model is as follows. Phosphorylation of p47phox leads to a conformational change allowing its interaction with p22phox(327, 843). It is thought that p47phox organizes the translocation
[page 250]
FIG. 3. Assembly of the phagocyte NADPH oxidase NOX2. [...] In resting neutrophil granulocytes, NOX2 and p22phox are found primarily in the membrane of intracellular vesicles. They exist in close association, costabilizing one another. Upon activation, there is an exchange of GDP for GTP on Rac leading to its activation. Phosphorylation of the cytosolic p47phox subunit leads to conformational changes allowing interaction with p22phox. The movement of p47phox brings with it the other cytoplasmic subunits, p67phox and p40phox, to form the active NOX2 enzyme complex. Once activated, there is a fusion of NOX2-containing vesicles with the plasma membrane or the phagosomal membrane. The active enzyme complex transports electrons from cytoplasmic NADPH to extracellular or phagosomal oxygen to generate superoxide (O2-). of other cytosolic factors, hence its designation as “organizer subunit.” The localization of p47phox to the membrane brings the “activator subunit” p67phox into contact with NOX2 (342) and also brings the small subunit p40phox to the complex. Finally, the GTPase Rac interacts with NOX2 via a two-step mechanism involving an initial direct interaction with NOX2 (214), followed by a subsequent interaction with p67phox (476, 508). Once assembled, the complex is active and generates superoxide by transferring an electron from NADPH in the cytosol to oxygen on the luminal or extracellular space. [...] |
The source is given once, but it is not clear that so much text and in particular the illustration are taken from it. The image is taken via copy-paste without reference. The source given, Roos et al. (2003), does not contain any of the copied material. |
|
| [19.] Arc/Fragment 028 01 - Diskussion Bearbeitet: 25. February 2014, 22:24 WiseWoman Erstellt: 21. February 2014, 22:45 (Graf Isolan) | Arc, BauernOpfer, Fragment, Gesichtet, Risau 1997, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 28, Zeilen: 1-15, legend to fig. 5 |
Quelle: Risau 1997 Seite(n): 671, Zeilen: 1; left col. 1-12.17-22; legend to fig. 1 |
|---|---|
| Chapter II: Mechanisms of vascular growth
When Leonardo da Vinci first speculated about the heart and the circulatory system, he could only analyse the mechanism of organ formation by analogy. So he suggested that the vasculature developed like a tree from a seed (the heart) by sprouting roots (the liver capillary network) and a trunk with major branches (the aorta and arteries, Fig. 5).
Fig. 5 – Analogy between the botanic and vascular tree as drawn by Leonardo da Vinci (taken from “The anatomy of man: the cardiovascular system (1508)). The drawing is annotated in his cryptic mirror-writing, for example the big seed of the left plant labelled “core” (heart) is indicated by the arrowhead71. Although sprouting of vessels is indeed a principal mechanism of the blood vessel formation termed angiogenesis, other mechanisms are now known. Rather than sprouting from the heart, the vascular system is laid down before the heart starts beating. Conversely, adult blood vessels generally form from pre-existing vessels in direct response to tissue demands74. II.1 Vasculogenesis and Angiogenesis The early vascular plexus forms from the mesoderm by differentiation of angioblasts (vascular endothelial cells that have not yet formed a lumen), which subsequently generate primitive blood vessels. The molecular mechanisms responsible for this process, termed vasculogenesis74, are starting to emerge (Fig. 6). 71. Haghighat A WD, Whalin MK, Cowan DP, Taylor WR. . Granulocyte colonystimulating factor and granulocyte macrophage colony-stimulating factor exacerbate atherosclerosis in apolipoprotein E-deficient mice. Circulation. 2007;115:2049-2054. 74. Risau W. Mechanisms of angiogenesis. Nature. 1997;386:671-674. |
Mechanisms of angiogenesis
[...] When Leonardo da Vinci first speculated about the heart and circulatory system (see Nature 380, 9; 1996), he could only analyse the mechanisms of organ formation by analogy, so he suggested that the vasculature developed like a tree from a seed (the heart) by sprouting roots (the liver capillary meshwork) and a trunk with major branches (the aorta and arteries) (Fig. 1). Although sprouting of blood vessels is indeed a principal mechanism of blood vessel formation, termed angiogenesis, other mechanisms are now known. Rather than sprouting from the heart, the vascular system is laid down before the heart starts beating. Conversely, adult blood vessels generally form from pre-existing vessels in direct response to tissue demands. [...] Vasculogenesis The early vascular plexus forms from mesoderm by differentiation of angioblasts (vascular endothelial cells that have not yet formed a lumen), which subsequently generate primitive blood vessels. The molecular mechanisms responsible for this process, termed vasculogenesis1, are emerging (Fig. 2).
Figure 1 Analogy between the botanic and the vascular tree as drawn by Leonardo da Vinci (taken from ‘the anatomy of man: the cardiovascular system’ (ca. 1508)). The drawing is annotated by notes in his trademark cryptic mirror-writing, for example the big seed of the left plant labelled ‘core’ (heart) is indicated by the arrowhead. 1. Risau, W. & Flamme, I. Vasculogenesis. Annu. Rev. Cell Dev. Biol. 11, 73-91 (1995). |


Although the source is given (in passing) nothing has been marked as a citation. The copied text is mostly taken verbatim. Figure 5 and its legend are also taken from Risau (1997), this time without the source being named. The named source Haghighat et al. (2007) does not contain any of the copied material. |
|
| [20.] Arc/Fragment 029 01 - Diskussion Bearbeitet: 26. February 2014, 22:26 Schumann Erstellt: 21. February 2014, 23:03 (Graf Isolan) | Arc, BauernOpfer, Fragment, Gesichtet, Risau 1997, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 29, Zeilen: 1-17, legend to fig. 6 |
Quelle: Risau 1997 Seite(n): 671, 672, Zeilen: 671: left col. 22-29 - right col. 1-9.23-25; 672: legend to fig. 2; left col. 2-7 |
|---|---|
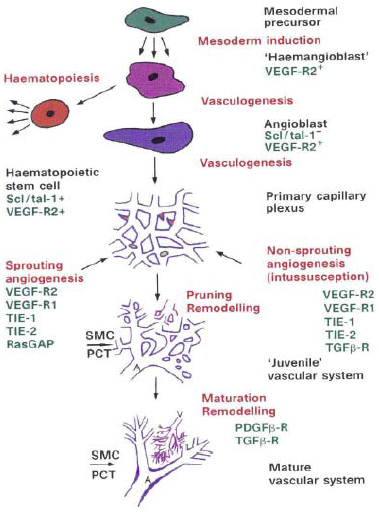
Fig. 6 – The processes (red labels), molecules (green labels) and appearances (black labels) involved in vascular development. Red tips in the primary capillary plexus represent sprouts, yellow circles represent splitting pillars. The hemangioblast as a bipotential precursor is still under debate and intermediate steps between some processes have been omitted. Remodelling and maturation is dependent on the tissue and organ context. It is schematized here from observations in the avian yolk-sac vascular system. A, arteriole; V, venule; SMC, smoothe muscle cells; PCT, pericytes71. Mesoderm inducing factors of the fibroblast growth factor family are crucial in inducing paraxial and lateral plate mesoderm to form angioblasts and hematopoietic cells. The existence of a bipotential precursor of these cell types (the so-called hemangioblast) is suggested by defects in both the hematopoietic and angioblastic lineages of embryos lacking VEGF-receptor 2 (VEGF-R2, also called Flk-1 and KDR in mice and humans, respectively). After differentiation, VEGF-R2 is downregulated in hematopoietic but not in endothelial cells. The other receptor for VEGF, VEGF-R1 (Flt-1), plays a later role, as mice lacking VEGF-R1 produce angioblasts, but their assembly into functional blood vessels is impaired75. VEGF acts in a paracrine manner as it is produced by the endoderm, whereas its receptors are expressed by mesoderm-derived angioblasts. Some angioblasts can migrate over long distances and form a vascular plexus at a site distant from the original location74. After the primary plexus is formed, more ECs are generated, which can form new capillaries by sprouting or by splitting from their vessel of origin in a process termed angiogenesis. There are at least two different types of angiogenesis: 1) nonsprouting or intussusception angiogenesis and 2) true sprouting of capillaries from preexisting vessels (Fig. 7)74. 71. Haghighat A WD, Whalin MK, Cowan DP, Taylor WR. . Granulocyte colonystimulating factor and granulocyte macrophage colony-stimulating factor exacerbate atherosclerosis in apolipoprotein E-deficient mice. Circulation. 2007;115:2049-2054. 74. Risau W. Mechanisms of angiogenesis. Nature. 1997;386:671-674. 75. Fong GH, Rossant J, Gertsenstein M, Breitman ML. Role of the Flt-1 receptor tyrosine kinase in regulating the assembly of vascular endothelium. Nature. 1995;376:66-70. |
[Page 671]
Mesoderm-inducing factors of the fibroblast growth factor family are crucial in inducing paraxial and lateral plate mesoderm to form angioblasts and haematopoietic cells. The existence of a bipotential precursor for these cell types (the so-called haemangioblast) is suggested by defects in both the haematopoietic and angioblastic lineages of embryos lacking vascular endothelial growth factor receptor-2 (VEGF-R2, also called Flk-1 and KDR in mice and humans, respectively; Fig. 2)4. But only haematopoietic stem cells are affected5,6 in embryos lacking the transcription factor Scl/tal-1. After differentiation, VEGF-R2 is downregulated in haematopoietic but not in endothelial cells. The other receptor for VEGF, VEGF-R1 (Flt-1), plays a role later, as mice lacking VEGF-R1 produce angioblasts, but their assembly into functional blood vessels is impaired7. VEGF itself acts in a paracrine manner as it is produced by the endoderm, whereas its receptors are expressed by mesoderm-derived angioblasts. [...] Rather than forming blood vessels where they originate, some angioblasts migrate over long distances12, forming a vascular plexus at a distant site (such as the perineural vascular plexus). [Page 672]
Figure 2 The processes (red labels), molecules (green labels) and appearances (black labels) involved in vascular development. Red tips in the primary capillary plexus represent sprouts, yellow circles represent splitting pillars (Fig. 3). The haemangioblast as a bipotential precursor is still hypothetical and intermediate steps between some processes have been omitted. Remodelling and maturation is dependent on the tissue and organ context. It is schematized here from observations in the avian yolk-sac vascular system. A, arteriole; V, venule; SMC, smooth muscle cells; PCT, pericytes. None of the figures is drawn to scale. [...] After the primary vascular plexus is formed, more endothelial cells are generated, which can form new capillaries by sprouting or by splitting from their vessel of origin in a process termed angiogenesis. There are at least two different types: true sprouting of capillaries from pre-existing vessels, and non-sprouting angiogenesis or intussusception (Figs 2 and 3). 4. Shalaby, F. et al. Failure o f blood-island formation and vasculogenesis in flk-1-deficient mice. Nature 376, 62-66 (1995). 5. Porcher, C. et al. The T cell leukemia oncoprotein Scl/Tal-1 is essential for development of all hematopoietic lineages. Cell 86, 47-57 (1996). 6. Robb, L. et al. The Scl gene product is required for the generation of all hematopoietic lineages in the adult mouse. EMBO J. 15, 4123-4129 (1996). 7. Fong, G. H., Rossant, J., Gertsenstein, M. & Breitman, M. L. Role of the fit-1 receptor tyrosine kinase in regulating the assembly of vascular endothelium. Nature 376, 66-70 (1995). 8. Carmeliet, P. et a l Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature 380, 435-439 (1996). 9. Ferrara, N. et al. Heterozygous embryonic lethality induced by targeted inactivation of the VEGF gene. Nature 380, 435-439 (1996). 12. Noden, D. M. in The development of the vascular system (eds Feinberg, R. N., Sherer, G. K. & Auerbach, R.) 1-24 (Karger, Basel, 1991). |

Although the source is given (in passing) nothing has been marked as a citation. The material is mostly copied verbatim. Figure 6 and its legend are also taken from Risau (1997), without the source being named. The source given Haghighat et al. (2007) does not contain any of the copied material. |
|
| [21.] Arc/Fragment 030 01 - Diskussion Bearbeitet: 26. February 2014, 22:35 Schumann Erstellt: 24. February 2014, 16:32 (Hindemith) | Arc, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Ten Dijke Arthur 2007, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 30, Zeilen: 1-5 |
Quelle: Ten Dijke Arthur 2007 Seite(n): 863, Zeilen: figure caption |
|---|---|
| In sprouting angiogenesis, endothelial cells proliferate behind the tip cell of a growing branch in response to cytokines, such as VEGF, and lumens can form by vacuole fusion. Both forms of angiogenesis require the recruitment of smooth muscle cells (SMCs) to stabilize the nascent vessels.
Fig. 7 – Representation of vasculogenesis and two types of angiogenesis: intussusceptive and sprouting angiogenesis. Vasculogenesis involves the differentiation of endothelial cells (ECs) from precursor angioblast cells to form a primitive plexus of capillaries, which remodel and grow by angiogenesis. Intussusceptive angiogenesis involves the splitting and growing of vessels in situ. Vessel splitting occurs by the formation of translumen pillars (arrowheads) but the molecular mechanisms are not well understood74. 74. Risau W. Mechanisms of angiogenesis. Nature. 1997;386:671-674. |
Figure 3 TGFβ signalling in vasculogenesis and angiogenesis. Vasculogenesis and two types of angiogenesis are shown: intussusceptive and sprouting angiogenesis (BOX 1). Vasculogenesis involves the differentiation of endothelial cells (ECs) from precursor angioblast cells to form a primitive plexus of capillaries, which remodel and grow by angiogenesis. Intussusceptive angiogenesis involves the splitting and growing of vessels in situ in a metabolically efficient manner, and is found, for example, in the developing yolk sac and lung. Vessel splitting occurs by the formation of translumen pillars (arrowheads) but the molecular mechanisms are not well understood. In sprouting angiogenesis, endothelial cells proliferate behind the tip cell of a growing branch in response to cytokines such as vascular endothelial growth factor (VEGF) and lumens can form by vacuole fusion. Both forms of angiogenesis require the recruitment of smooth muscle cells (SMCs) to stabilize the nascent vessels. |
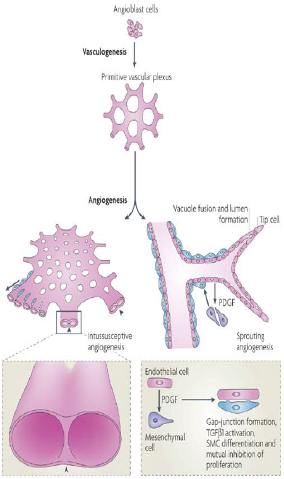
The source is not mentioned here. Risau (1997) does not contain the copied material. |
|
| [22.] Arc/Fragment 031 22 - Diskussion Bearbeitet: 27. February 2014, 00:48 Hindemith Erstellt: 23. February 2014, 19:57 (Hindemith) | Arc, BauernOpfer, Fragment, Gesichtet, Koerselman et al 2003, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 31, Zeilen: 22-29 |
Quelle: Koerselman et al 2003 Seite(n): 2507, Zeilen: l.col: 24-29, r.col:23-27 |
|---|---|
| Coronary collaterals, or “natural bypasses,” are anastomotic connections without an interposed capillary bed either between portions of the same coronary artery or between different coronary arteries83, potentially offering an important alternative source of blood supply when the original vessel fails to provide sufficient blood84. Collateral growth (commonly termed arteriogenesis) refers to the transformation of pre-existing (collateral) arterioles into functional (muscular) collateral arteries, as a thick muscular coat is added, concomitant with the acquisition of viscoelastic and vasomotor properties76.
76. Carmeliet P, Conway EM. Growing better blood vessels. Nat Biotechnol. 2001;19:1019-1020. 83. Koerselman J vdGY [sic], de Jaegere PP, Grobbee DE. Coronary collaterals: an important and underexposed aspect of coronary artery disease. Circulation. . 2003 107:2507-2511. 84. Sasayama S, Fujita M. Recent insights into coronary collateral circulation. Circulation. 1992;85:1197-1204. |
Coronary collaterals, or “natural bypasses,” are anastomotic connections without an intervening capillary bed between portions of the same coronary artery and between different coronary arteries (Figure 1).1 Collateral circulation potentially offers an important alternative source of blood supply when the original vessel fails to provide sufficient blood.2 [...]
Finally, arteriogenesis refers to the transformation of preexisting (collateral) arterioles into functional (muscular) collateral arteries, as a thick muscular coat is added, concomitant with acquisition of viscoelastic and vasomotor properties.10 1. Popma JJ, Bittl J. Coronary angiography and intravascular ultrasonography. In: Braunwald E, Zipes DP, Libby P, eds. Heart Disease: A Textbook of Cardiovascular Medicine. Philadelphia: W.B. Saunders Company; 2001:387–418. 2. Sasayama S, Fujita M. Recent insights into coronary collateral circulation. Circulation. 1992;85:1197–1204. 10. Conway EM, Collen D, Carmeliet P. Molecular mechanisms of blood vessel growth. Cardiovasc Res. 2001;49:507–521. |
The source is given, but it is not clear to the reader that text has been copied 1-to-1 and that the copied text continues after the reference to Koerselman et al. (2003). The reference to Sasayama & Fujita (1992) is also taken from the source. None of the copied text can be found either in Sasayama & Fujita (1992) or in Carmeliet & Conway (2001). |
|
| [23.] Arc/Fragment 032 10 - Diskussion Bearbeitet: 26. February 2014, 22:31 Schumann Erstellt: 24. February 2014, 18:28 (Hindemith) | Arc, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Simons Ware 2003, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 32, Zeilen: 10-13 |
Quelle: Simons Ware 2003 Seite(n): 2, 3, 4, Zeilen: 2: r.col: last lines; 3: l.col: 1; 4: figure |
|---|---|

Fig. 8 – A: Collateral vessels can develop around the site of coronary occlusion, via remodelling of pre-existing vessels (arteriogenesis) that gradually enlarge to the point at which they can carry the bulk of blood flow84. [...] While the development of vasculature in the course of embryonic growth is beginning to be well understood, comparatively little information is available about the processes responsible for coronary collateral growth. 84. Sasayama S, Fujita M. Recent insights into coronary collateral circulation. Circulation. 1992;85:1197-1204. |
Whereas the development of vasculature in the course of embryonic growth is beginning to be well understood, comparatively little information is available about the processes responsible for vessel growth and maintenance
[page 3] in mature adult tissues. [page 4]
Figure 2 Arteriogenesis. Collateral vessels (arteriogenesis) can develop around the site of coronary occlusion. Although the exact mechanism is not clear, there are two distinct possibilities. One option (a) for arteriogenesis is to proceed via remodelling of pre-existing vessels that gradually enlarge to the point at which they can carry the bulk of blood flow. |
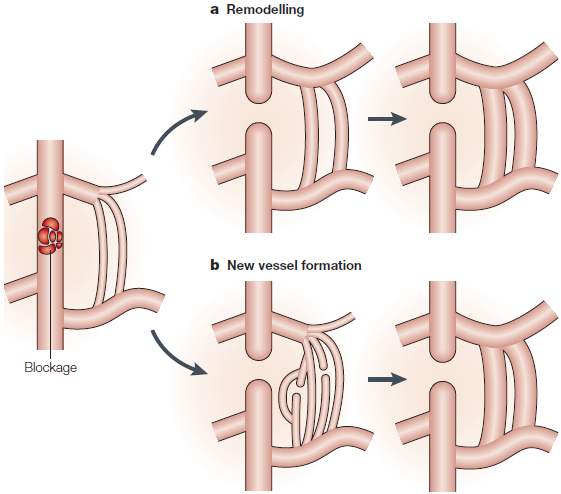
The source is not given. The copied material cannot be found in the given reference Sasayama & Fujita (1992). Note that the part B of figure 8 has been taken from another publication: Arc/Fragment_032_11 |
|
| [24.] Arc/Fragment 032 11 - Diskussion Bearbeitet: 27. February 2014, 00:23 Hindemith Erstellt: 23. February 2014, 21:07 (Hindemith) | Arc, Fragment, Gesichtet, Koerselman et al 2003, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 32, Zeilen: 10-10 |
Quelle: Koerselman et al 2003 Seite(n): 2508, Zeilen: top of page |
|---|---|

Fig. 8 – A: [...] B: Left anterior oblique view of the right coronary arteriogram. The left circumflex coronary artery (LCX) is proximally occluded and fills completely by means of collateral circulation from the right coronary artery (RCA)81. 81. Bloor CM, Liebow AA. Coronary Collateral Circulation. Am J Cardiol. 1965;16:238-252. |
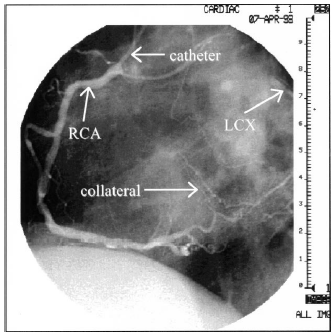
Figure 1. Left anterior oblique view of the right coronary arteriogram. The left circumflex coronary artery (LCX) is proximally occluded and fills completely by means of collateral circulation from the right coronary artery (RCA). Image courtesy of the Department of Cardiology at the Heronimus Bosch Hospital, Den Bosch, the Netherlands. |
The given source has not been checked. However, the copied image contains the date "7-Apr-93" (or "7-Apr-98"), such that it cannot have been taken from a publication of 1965. The source is mentioned on the previous page, but without any indication that this image might have been taken from it. Note that the part A of figure 8 has been taken from a different publication: Arc/Fragment_032_10 |
|
| [25.] Arc/Fragment 033 25 - Diskussion Bearbeitet: 26. February 2014, 22:40 Schumann Erstellt: 23. February 2014, 18:10 (Hindemith) | Arc, BauernOpfer, Fragment, Gesichtet, Heil Schaper 2004, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 33, Zeilen: 25-29 |
Quelle: Heil Schaper 2004 Seite(n): 450, Zeilen: r.col: 14ff |
|---|---|
| [The role of shear stress in collateral growth has been discussed extensively by Wolfgang Schaper’s research group and others91, 92. Their model is based on the following concept (Fig. 9):] After occlusion of an artery, the pressure in its distal stump falls to very low levels. When a vascular connection like a pre-existent collateral anastomoses [sic] exists between the high-pressure (proximal to the occlusion) and the low pressure (distal from the occlusion) region, a steep pressure gradient develops which increases blood flow through these connections.
91. Helisch A, Wagner S, Khan N, Drinane M, Wolfram S, Heil M, Ziegelhoeffer T, Brandt U, Pearlman JD, Swartz HM, Schaper W. Impact of mouse strain differences in innate hindlimb collateral vasculature. Arterioscler Thromb Vasc Biol. 2006;26:520-526. 92. Cai Z, Zhong H, Bosch-Marce M, Fox-Talbot K, Wang L, Wei C, Trush MA, Semenza GL. Complete loss of ischaemic preconditioning-induced cardioprotection in mice with partial deficiency of HIF-1{alpha}. Cardiovasc Res. 2008;77:463-470. |
On occlusion of an artery, the pressure in its distal stump falls to very low levels. When a vascular connection like a preexistent collateral anastomosis exists between the high-pressure proximal to the occlusion with the low-pressure region distal from the occlusion, a steep pressure gradient develops which increases blood flow through these connections (Figure 1). |
To be continued on the next page: Arc/Fragment 034 01 The authors of the source are mentioned, the source itself is not. The given sources Cai et al. (2009) and Helisch et al. (2006) do not contain the copied text. Nothing is marked as a quotation. |
|
| [26.] Arc/Fragment 034 01 - Diskussion Bearbeitet: 25. February 2014, 23:25 WiseWoman Erstellt: 23. February 2014, 17:22 (Hindemith) | Arc, BauernOpfer, Fragment, Gesichtet, Heil Schaper 2004, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 34, Zeilen: 1-11 |
Quelle: Heil Schaper 2004 Seite(n): 450, 451, Zeilen: 450: r.col: 1ff; 451: l.col: 1ff |
|---|---|
 Fig. 9 – Diagram of blood flow changes after occlusion of the femoral artery in the rabbit hind limb. After occlusion of the femoral artery (X), blood flow (arrows) follows the gradient between pressures proximal to the occlusion site and very low distal pressures, recruting pre-existing collateral arterioles89. According to Heil and Schaper91, the vessel wall is now exposed to pronounced mechanical forces: increased blood flow directly augments fluid shear stress (FSS), i.e., the viscous drag that flowing blood exerts on the endothelial lining. By assuming Newtonian fluid dynamics, FSS can be estimated using the following equation:  The authors demonstrate that blood flow (Q) will directly result in increased FSS (&tao;). The wall of the collateral arteriole is influenced by pressure-related forces like longitudinal-, circumferential- and radial-wall stresses. The distension of the vessel wall, structurally weakened by intravascular pressure, increases the circumferential wall stress (CWS). FSS is a relatively weak force compared with CWS, which is 106 times higher, posing the hypothesis that FSS can only act in concert with pressure-dependent forces91. 89. Gho BC, Schoemaker RG, van den Doel MA, Duncker DJ, Verdouw PD. Myocardial protection by brief ischemia in noncardiac tissue. Circulation. 1996;94:2193-2200. 91. Helisch A, Wagner S, Khan N, Drinane M, Wolfram S, Heil M, Ziegelhoeffer T, Brandt U, Pearlman JD, Swartz HM, Schaper W. Impact of mouse strain differences in innate hindlimb collateral vasculature. Arterioscler Thromb Vasc Biol. 2006;26:520-526. |
 Figure 1. Diagram of blood flow changes after occlusion of the femoral artery in the rabbit hind limb. After occlusion of the femoral artery (X), blood flow (arrows) follows the gradient between high pressures proximal to the occlusion site and very low distal pressures. Preexisting collateral arterioles are recruited. [...] [...] Hence, the collateral vessel wall is now exposed to various pronounced mechanical forces: increased blood flow directly augments fluid shear stress (FSS), ie, the viscous drag that flowing blood exerts on the endothelial lining. Assuming Newtonian fluid dynamics, FSS can be estimated using the following equation: 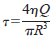 The equation that already includes blood viscosity (η) and the internal radius of a vessel (R), demonstrates that increased blood flow (Q) will directly result in increased FSS (&tao;).8 Furthermore, the wall of the collateral arteriole is influenced by pressure-related forces like longitudinal-, circumferential-, and radial wall stresses. The distention of the vessel wall, structurally weakened by matrix digestion, by the intravascular pressure, increases the circumferential wall stress, a known activator of smooth muscle cells (SMCs) proliferation.9 In contrast, FSS is a relatively weak force, more than two orders of magnitude lower than the pressure-derived forces [page 451] acting on the arterial wall. The difference is so impressive that other authors questioned the morphogenic force of FSS or posed the hypothesis that FSS can act only in concert with the pressure-dependent forces.9 8. Cox R. Physiology and hemodynamics of the macrocirculation. In: Stehbens W, eds. Hemodynamics and the Blood Vessel Wall. Springfield, Ill: Charles C. Thomas; 1979:75–156. 9. Scheel KW, Fitzgerald EM, Martin RO, Larsen RA. The possible role of mechanical stresses on coronary collateral development during gradual coronary occlusion. In: Schaper W, eds. The Pathophysiology of Myocardial Perfusion. Amsterdam: Elsevier/North-Holland; 1979:489–518. |
The source is mentioned in the text (also on the previous page: Arc/Fragment 033 25), but it is not clear that the text as well as the diagram is taken 1-to-1 from the source, which is not listed in the bibliography. Note also that in the given sources Gho et al. (1996) and Helisch et al. (2006) the copied material cannot be found. |
|
| [27.] Arc/Fragment 036 12 - Diskussion Bearbeitet: 25. February 2014, 21:58 Hindemith Erstellt: 23. February 2014, 07:45 (Hindemith) | Arc, Bedard Krause 2007, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 36, Zeilen: 12-14 |
Quelle: Bedard Krause 2007 Seite(n): 246, Zeilen: 29-33 |
|---|---|
| Through such interactions, ROS may irreversibly destroy or alter the function of the target molecule and consequently, ROS have been identified as major contributors to damage in biological organisms. | Through such interactions, ROS may irreversibly destroy or alter the function of the target molecule. Consequently, ROS have been increasingly identified as major contributors to damage in biological organisms. |
A reference to the source is missing. Note that this phrase has been used before here: Arc/Fragment 022 01 |
|
| [28.] Arc/Fragment 037 01 - Diskussion Bearbeitet: 6. March 2014, 09:16 Singulus Erstellt: 23. February 2014, 15:14 (Hindemith) | Arc, BauernOpfer, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Ushio-Fukai 2006 |
|
|
|
| Untersuchte Arbeit: Seite: 37, Zeilen: 0-1 |
Quelle: Ushio-Fukai 2006 Seite(n): 231, Zeilen: 1ff |
|---|---|
| [In Fig. 10, Ushio-Fukai describes important ROS-dependent genes and proteins associated with anigiogenesis, like urokinase plasminogen activator (uPA), plasminogen activator inhibitor-1 (PAI-1) and metalloproteinases (MMPs).]
Fig. 10 - Role of ROS derived from NADPH oxidase in the induction of transcription factors and genes involved in angiogenesis. Ischemia/hypoxia stimulates VEGF induction and NADPH oxidase, thereby promoting activation of redox signalling events and induction of redox sensitive transcription factors and genes involved in angiogenesis. ROS derived from NADPH oxidase also upregulate mitochondrial SOD (MnSOD), increasing mitochondrial H2O2 production, which could represent a feed-forward mechanism by which ROS-triggered H2O2 formation plays an important role in angiogenesis101. 101. Tuttle JL, Sanders BM, Burkhart HM, Fath SW, Kerr KA, Watson WC, Herring BP, Dalsing MC, Unthank JL. Impaired collateral artery development in spontaneously hypertensive rats. Microcirculation. 2002;9:343-351. |
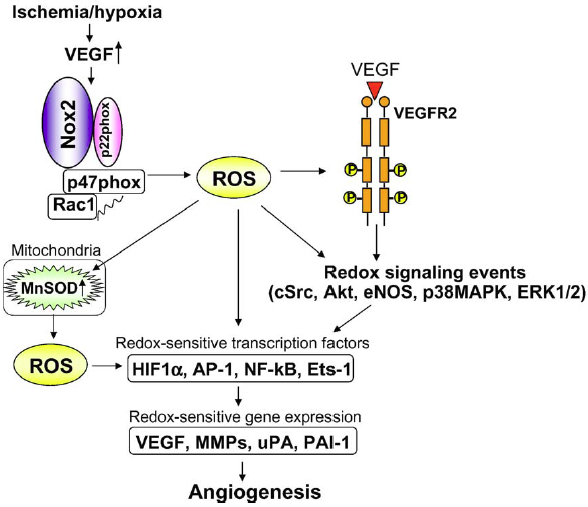
Fig. 4. Role of ROS derived from NADPH oxidase in induction of transcription factors and genes involved in angiogenesis. Ischemia/hypoxia stimulates VEGF induction and NADPH oxidase, thereby promoting activation of redox signaling events and induction of redox sensitive transcription factors and genes involved in angiogenesis. ROS derived from NADPH oxidase also upregulate MnSOD which increases mitochondrial H2O2 production, which could represent a feed-forward mechanism by which ROS-triggered H2O2 formation plays an important role in angiogenesis. |
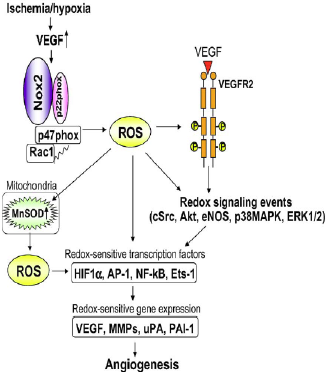
On the previous page it is mentioned that "In Fig. 10, Ushio-Fukai describes important ...". However, there is no reference to a publication by Usio-Fukai to be found in the list of references, and it is also not clear that the caption of figure 10 is also taken from that source. In the given source Tuttle et al. (2002) none of the copied material can be found. |
|
| [29.] Arc/Fragment 037 11 - Diskussion Bearbeitet: 26. February 2014, 00:43 Hindemith Erstellt: 25. February 2014, 01:29 (Hindemith) | Arc, Fragment, Gesichtet, Maulik et al 1998, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 37, Zeilen: 11-16 |
Quelle: Maulik et al 1998 Seite(n): 365, Zeilen: l.col: 33-40 |
|---|---|
| Tissue such as the myocardium can be adapted to ischemic stress by repeatedly subjecting it to short terms of reversible ischemia, each followed by another short duration of reperfusion108, 109. This phenomenon causes the production of oxidative stress, leading to the induction of gene expression, which is subsequently translated into the development of beneficial proteins responsible for the heart’s defence97.
97. Murphy E, Steenbergen C. Mechanisms Underlying Acute Protection From Cardiac Ischemia-Reperfusion Injury. Physiol. Rev. 2008;88:581-609. 108. Maulik N, Yoshida T, Zu YL, Sato M, Banerjee A, Das DK. Ischemic preconditioning triggers tyrosine kinase signaling: a potential role for MAPKAP kinase 2. Am J Physiol. 1998;275:H1857-1864. 109. Maulik N, Yoshida T, Engelman RM, Deaton D, Flack JE, 3rd, Rousou JA, Das DK. Ischemic preconditioning attenuates apoptotic cell death associated with ischemia/reperfusion. Mol Cell Biochem. 1998;186:139-145. |
Mammalian heart can be adapted to ischemia by repeatedly subjecting it to short-term reversible ischemia each followed by another short duration of reperfusion [1,2]. This phenomenon, known as ischemic adaptation, causes the production of oxidative stress leading to the induction of gene expression which is subsequently translated into the development of several stress-related proteins responsible for the heart's defense [3].
[1] Reimer, A., vander Heide, R.S. and Jennings, R.B. (1994) Ann. NY Acad. Sci. 723, 99-115. [2] Banerjee, A., Locke-Winter, C., Rogers, K.B., Mitchell, M.B., Bensard, D.D., Brew, E.C., Cairns, C.B. and Harken, A.H. (1993) Circ. Res. 73, 656-670. [3] Das, D.K., Maulik, N. and Moraru, I.I. (1995) J. Mol. Cell. Cardiol. 27, 181-193. |
The source is not given (the two papers by Maulik et al. given are different to the source here documented). The papers referenced under 97, 108 and 109 do not contain the copied text. |
|
| [30.] Arc/Fragment 038 01 - Diskussion Bearbeitet: 26. February 2014, 23:46 Hindemith Erstellt: 25. February 2014, 17:07 (Hindemith) | Arc, Fragment, Gesichtet, Liu et al 2008, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 38, Zeilen: 1-11 |
Quelle: Liu et al 2008 Seite(n): 1, 2, Zeilen: 1: 33-39 - 2: 1-3,9-11 |
|---|---|
| [It was later found that the generation of ROS was a result of the opening of mitochondrial ATP-sensitive potassium channels (mKATP), acting as second messengers in a pathway that] ended in the activation of protein kinase C (PKC), which would then trigger the preconditioned state111. In those studies, ROS were recognized as part of the trigger pathway that preconditions the heart prior to the onset of ischemia. It was discovered that IPC actually exerts its protective effect in the first minutes of reperfusion112, presumably by inhibiting the formation of mitochondrial permeability transition pores (mPTP)113. It was also found that redox signalling is part of the pathway that mediates IPC protection at the time of reperfusion, meaning that ROS generation was required for IPC to be protective114. It is unknown how redox signalling acts to mediate protection at reperfusion115, but a number of signal transduction components have been identified in IPC’s mediator pathway, including phosphatidylinositol (PI) 3-kinase and extracellular signal-regulated kinase (ERK)112.
111. Pain T, Yang XM, Critz SD, Yue Y, Nakano A, Liu GS, Heusch G, Cohen MV, Downey JM. Opening of mitochondrial K(ATP) channels triggers the preconditioned state by generating free radicals. Circ Res. 2000;87:460-466. 112. Hausenloy DJ, Tsang A, Mocanu MM, Yellon DM. Ischemic preconditioning protects by activating prosurvival kinases at reperfusion. Am J Physiol Heart Circ Physiol. 2005;288:H971-976. 113. Hausenloy DJ, Yellon DM, Mani-Babu S, Duchen MR. Preconditioning protects by inhibiting the mitochondrial permeability transition. Am J Physiol Heart Circ Physiol. 2004;287:H841-849. 114. Penna C, Rastaldo R, Mancardi D, Raimondo S, Cappello S, Gattullo D, Losano G, Pagliaro P. Post-conditioning induced cardioprotection requires signaling through a redox-sensitive mechanism, mitochondrial ATP-sensitive K+ channel and protein kinase C activation. Basic Res Cardiol. 2006;101:180-189. 115. Liu F, Patient R. Genome-Wide Analysis of the Zebrafish ETS Family Identifies Three Genes Required for Hemangioblast Differentiation or Angiogenesis. Circ Res. 2008:CIRCRESAHA.108.179713. |
[page 1]
Later it was found that ROS were generated as a result of opening of mitochondrial ATP-sensitive potassium channels (mKATP) and acted as second messengers in a pathway ending in activation of PKC which triggered the preconditioned state (20). In those studies ROS were recognized as part of the trigger pathway that preconditions the heart prior to the onset of the index ischemia. Several years ago it was discovered that IPC actually exerts its protective effect in the first minutes of reperfusion (9) presumably by inhibiting the formation of mitochondrial permeability [page 2] transition pores (mPTP) (11). Then it was found that redox signaling is part of the pathway that mediates IPC's protection at the time of reperfusion as well. Penna et al. (21) reported that ROS generation was required for ischemic postconditioning to be protective. [...] It is unknown how redox signaling acts to mediate protection at reperfusion. A number of signal transduction components have been identified in IPC's mediator pathway including phosphatidylinositol (PI) 3-kinase and extracellular signal-regulated kinase (ERK) (9). 9. Hausenloy DJ, Tsang A, Mocanu MM, Yellon DM. Ischemic preconditioning protects by activating prosurvival kinases at reperfusion. Am J Physiol 2005;288:H971–H976. 11. Hausenloy DJ, Yellon DM, Mani-Babu S, Duchen MR. Preconditioning protects by inhibiting the mitochondrial permeability transition. Am J Physiol 2004;287:H841–H849. 20. Pain T, Yang X-M, Critz SD, Yue Y, Nakano A, Liu GS, Heusch G, Cohen MV, Downey JM. Opening of mitochondrial KATP channels triggers the preconditioned state by generating free radicals. Circ Res 2000;87:460–466. [PubMed: 10988237] 21. Penna C, Rastaldo R, Mancardi D, Raimondo S, Cappello S, Gattullo D, Losano G, Pagliaro P. Postconditioning induced cardioprotection requires signaling through a redox-sensitive mechanism, mitochondrial ATP-sensitive K+ channel and protein kinase C activation. Basic Res Cardiol 2006;101:180–189. [PubMed: 16450075] |
The source is not mentioned. The copied text cannot be found in Liu & Patient (2008). |
|
| [31.] Arc/Fragment 040 03 - Diskussion Bearbeitet: 26. February 2014, 23:06 Guckar Erstellt: 23. February 2014, 14:21 (Hindemith) | Arc, BauernOpfer, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Toyota et al 2005 |
|
|
|
| Untersuchte Arbeit: Seite: 40, Zeilen: 3-32 |
Quelle: Toyota et al 2005 Seite(n): 2109, Zeilen: l.col: 10-46 |
|---|---|
| Male Sprague-Dawley rats (3–4 mo old, 300–350 g) were used for chronic (5 days) implantation of a pneumatic occluder over the left anterior descending coronary artery (LAD; as described by Toyota et al.96) to produce repetitive ischemia (RI). Briefly, for surgery, rats were premedicated (ketamine 50 mg/mL plus acepromazine 2.5 mg/mL plus torbuterol 2.0 mg/mL, 0.2 mL/100 g body weight, i.p.) and intubated. Oral intubation (I6-G polyethylene tubing) was done under direct observation of the vocal cords with an otoscope. General anesthesia was introduced and maintained by sevoflurane inhalation (1.0% to 2.0%, with 100% oxygen). Body temperature was controlled at 37°C by an electric heating table. Surgery was performed using aseptic technique. The animal was initially placed on its dorsal side, and after a neck incision, the right carotid artery was isolated, and a PE-50 catheter filled with heparin (10 U/mL)-saline was inserted. This tubing was used for monitoring of systemic hemodynamics, sampling arterial blood, and maintaining the blood volume. Blood pH, PaO2, PaCO2, and systemic hemodynamics were maintained within physiological ranges throughout the surgery. The animal was repositioned on its right side, and the heart was exposed by left thoracotomy. A mini-pneumatic snare occluder (see the Mini- Pneumatic Snare Occluder section for details) was implanted around the mid to proximal LAD. Confirmation that the occluder was functional, i.e., producing myocardial ischemia, was determined initially by observation of blanching and hypokinesis of the left ventricle (LV) during inflation. Rats were randomly divided into 3 groups based on the type of measurement: coronary blood flow (CBF) (neutron activated microspheres), oxidative stress analysis (Dihydroethidium, DHE) or vascular imaging (micro-CT). CBF was measured during coronary occlusion to determine flows to the normal and collateral-dependent zones (see the Microsphere measurements of myocardial and collateral-dependent blood flow and the Coronary Microvascular imaging with Cryomicrotome sections). After instrumentation and measurements, the chest was closed under positive end-expiratory pressure, and the thoracic cavity was evacuated of air. The occluder was tunnelled subcutaneously and exteriorized between the scapulae. This catheter was protected by a stainless steel spring coil connected to a ring that was secured subcutaneously between the scapulae.
96. Toyota E, Warltier DC, Brock T, Ritman E, Kolz C, O'Malley P, Rocic P, Focardi M, Chilian WM. Vascular endothelial growth factor is required for coronary collateral growth in the rat. Circulation. 2005;112:2108-2113. |
Male Wistar rats (290- to 360-g body weight; n=50) were used for experiments (preparations were successful in 39 animals). For surgery, rats were premedicated (ketamine 50 mg/mL plus acepromazine 2.5 mg/mL plus torbuterol 2.0 mg/mL, 0.2 mL/100 g body weight IP) and intubated. Oral intubation (I6-G polyethylene tubing) was done under direct observation of the vocal cords with an otoscope. General anesthesia was introduced and maintained by sevoflurane inhalation (1.0% to 2.0%, with 100% oxygen). Body temperature was controlled at 37°C by an electric heating table. Surgery was performed using aseptic technique. The animal was initially placed on its dorsal side, and after a neck incision, the right carotid artery was isolated, and a PE-50 catheter filled with heparin (10 U/mL)-saline was inserted. This tubing was used for monitoring of systemic hemodynamics, sampling arterial blood, and maintaining the blood volume. Blood pH, PaO2, PaCO2, and systemic hemodynamics were maintained within physiological ranges throughout the surgery. The animal was repositioned on its right side, and the heart was exposed by left thoracotomy. A mini-pneumatic snare occluder (see the Mini-Pneumatic Snare Occluder section for details) was implanted around the mid to proximal left anterior descending coronary artery (LAD). Confirmation that the occluder was functional, ie, producing myocardial ischemia, was determined initially by observation of blanching and hypokinesis of the left ventricle (LV) during inflation. Rats were randomly divided into 2 groups based on the type of measurement: coronary blood flow (CBF) (radioactive microspheres, n=20) or vascular imaging (micro-CT, n=13). CBF was measured during coronary occlusion to determine flows to the normal and collateral-dependent zones (see the Measurement of Coronary Blood Flow and the Coronary Vascular Imaging With Micro-CT sections). After instrumentation and measurements, the chest was closed under positive end-expiratory pressure, and the thoracic cavity was evacuated of air. An intraperitoneal catheter (PE-50) for drug administration was inserted, and this catheter and the occluder were tunneled subcutaneously and exteriorized between the scapulae. These catheters were protected by a stainless steel spring coil connected to a ring that was secured subcutaneously between the scapulae. |
The source is given at the beginning of the section in a way that indicates that the here described method is similar to the one employed by the authors of the source. However, it is not clear to the reader that the description of the method is a largely literal copy of the source. |
|
| [32.] Arc/Fragment 041 01 - Diskussion Bearbeitet: 26. February 2014, 23:07 Guckar Erstellt: 23. February 2014, 08:42 (Hindemith) | Arc, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Toyota et al 2005, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 41, Zeilen: 1-22 |
Quelle: Toyota et al 2005 Seite(n): 2109, Zeilen: l.col: 46ff |
|---|---|
| [After the surgery, analgesic] (buprenorphine 0.05 mg/kg s.c.) and antibiotic (enrofloxacin 10 mg/kg s.c.) compounds were administered. Rats were observed in a recovery cage for 2 hours and then transferred to the animal care facility. For 3 days after the surgery, buprenorphine (0.5 mg/kg BID mixed in strawberry Jello) was taken orally for pain relief. On the fourth day after the surgery, the ischemic protocol was started (see the Experimental Protocol section). After 5 days of the experimental protocol, the rats were anesthetized and the chest was opened by mid thoracotomy. In CBF measurement or DHE analysis groups, the hearts were excised at the end of procedures, and the tissue was prepared for analyses.
3.2 Mini-Pneumatic Snare Occluder for Rat Heart A mini-pneumatic snare occluder consisting of a mini-balloon, sheath tubing, suture, and catheter (Fig. 11) was placed on the LAD of the rat heart. The balloon (7 mm long) is made of soft latex membrane and is sufficiently pliable to give negligible physical force on the coronary vessels during balloon deflation. The balloon is mounted within an umbrella sheath (3.2 or 4.8 mm in diameter, 12 mm in length; protects the balloon from fibrous infiltration). Prolene (5– 0) is passed around the LAD and attached to the sheath, securing the occluder to the heart, so that myocardial ischemia is produced by balloon inflation. Inflation volume is small (0.2 to 0.25 mL air), but occlusion occurs by 2 physical actions: “crimping” the LAD toward upward/outside and compressing the LAD by the inflated balloon/sheath. The balloon is connected to a catheter (PE-50) that is exteriorized. Balloon inflation and deflation are controlled from outside the rat cage.
Fig. 11 - Schematic diagram of the mini-pneumatic snare and its actions. Top: Cross-sectional and longitudinal views when the balloon is deflated. Bottom: Views during inflation. The artery is patent when the balloon is deflated, but during inflation, a snare situated underneath the artery is pulled “upward” during inflation, producing the coronary occlusion94. 94. Kappel A, Ronicke V, Damert A, Flamme I, Risau W, Breier G. Identification of Vascular Endothelial Growth Factor (VEGF) Receptor-2 (Flk-1) Promoter/Enhancer Sequences Sufficient for Angioblast and Endothelial Cell- Specific Transcription in Transgenic Mice. Blood. 1999;93:4284-4292. |
After the surgery, analgesic (buprenorphine 0.05 mg/kg SC) and antibiotic (enrofloxacin 10 mg/kg SC) were administered. Rats were observed in a recovery cage for 2 hours and then transferred to the animal care facility. For 3 days after the surgery, buprenorphine (0.5 mg/kg BID mixed in strawberry Jello) was taken orally for pain. On the fourth day after the surgery, ischemic protocol was started (see the Experimental Protocol section).
After 10 days of the experimental protocol, the rats were anesthetized, and the chest was opened by mid thoracotomy. In the micro-CT group, the hearts were immediately excised. In CBF measurement group, blood flow to the normal and collateraldependent zones during coronary occlusion was measured. The heart was excised at the end of measurements, and the tissue was prepared for analyses of radioactivity. Mini-Pneumatic Snare Occluder for Rat Heart We developed a mini-pneumatic snare occluder (patent application serial number: 11/071,617, E.T. and W.M.C.) consisting of a mini-balloon, sheath tubing, suture, and catheter (Figure 1). The balloon (7 mm long) is made of soft latex membrane and is sufficiently pliable to give negligible physical force on the coronary vessels during balloon deflation. The balloon is mounted within an umbrella sheath (3.2 or 4.8 mm in diameter, 12 mm in length; protects the balloon from fibrous infiltration). Prolene (5– 0) is passed around the LAD and attached to the sheath, securing the occluder to the heart, so that myocardial ischemia is produced by balloon inflation. Inflation volume is small (0.2 to 0.25 mL air), but occlusion occurs by 2 physical actions: “crimping” the LAD toward upward/outside and compressing the LAD by the inflated balloon/sheath. The balloon is connected to a catheter (PE-50) that is exteriorized. Balloon inflation and deflation are controlled from outside the rat cage.
Figure 1. Schematic of the mini-pneumatic snare and its actions. Top, Cross-sectional and longitudinal views when the balloon is deflated. Bottom, Views during inflation. The artery is patent when the balloon is deflated, but during inflation, a snare situated underneath the artery is pulled “upward” during inflation, producing the coronary occlusion. |

At the beginning of the previous page the source is mentioned, but without indication that the following two pages are taken from it. The given source Kappel et al. (1999) doesn't contain any of the copied material. |
|
| [33.] Arc/Fragment 042 16 - Diskussion Bearbeitet: 26. February 2014, 23:07 Guckar Erstellt: 23. February 2014, 12:32 (Hindemith) | Arc, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Toyota et al 2005, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 42, Zeilen: 16-29 |
Quelle: Toyota et al 2005 Seite(n): 2109, Zeilen: r.col: 13ff |
|---|---|
| CBF was measured with neutron activated microspheres (Biopal, 15 μm) before initiation of (after all the instruments had been implanted at the time of the initial surgery) and at the end of the repetitive occlusions (when the rats were anesthetized and the chest was open to mimic the conditions of the first measurement) to measure normal zone flows and flow in the developed collaterals. For the first measurement, neutron activated microspheres labelled with Samarium were mixed with fluorescent (FITC) microspheres (Invitrogen, 10 μm) to identify the collateral-dependent region as described below. For the second measurement, Gold labelled microspheres were used. The microspheres (5×105) were injected directly into the LV cavity via the LV apex during LAD occlusion with a 29-gauge insulin syringe over a 10-second period. During the course of the procedures, systemic pressure and heart rate were recorded (386- BIOS, American Megatrends Inc). The heart was excised and fresh LV was sliced along the short axis and observed with a dissecting microscope and fluorescent light source. The collateral-dependent area (ischemic zone, LAD region) was distinguished as the [area without fluorescent microspheres.] | CBF was measured with radioactive microspheres (Perkins Elmer; φ; 15 μm; 95Nb and 103Ru, ≈4.5x105) before initiation of (after all the instruments had been implanted at the time of the initial surgery) and at the end of the repetitive occlusions (when the rats were anesthetized and the chest was open to mimic the conditions of the first measurement) to measure normal zone flows, native collateral flow, and flow in the developed collaterals. For the first measurement, radioactive microspheres were mixed with fluorescent (FITC) microspheres (φ, 10 μm; ≈9x105, Fluoresbrite Yellow Green, Polysciences, Inc) to identify the collateral-dependent region as described below. For the second measurement, the other nuclidelabeled microspheres were used. The microspheres were agitated for 15 minutes, suspended in saline (total volume, 150 μL), and then injected directly into the LV cavity via the LV apex during LAD occlusion with a 29-gauge insulin syringe over a 10-second period. [...] During the course of the procedures, systemic pressure and heart rate were recorded (386- BIOS, American Megatrends Inc).
The heart was excised and fixed in 4% paraformaldehyde solution overnight. The fixed LV was sliced along the short axis and observed with a dissecting microscope and fluorescent light source (LT-9800, Lightools Research). The collateral-dependent area (LAD region) was distinguished as the area without fluorescent microspheres. |
The source is not given. |
|
| [34.] Arc/Fragment 043 01 - Diskussion Bearbeitet: 27. February 2014, 19:44 WiseWoman Erstellt: 23. February 2014, 13:29 (Hindemith) | Arc, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Toyota et al 2005, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 43, Zeilen: 1-5 |
Quelle: Toyota et al 2005 Seite(n): 2109, Zeilen: r.col: 40ff |
|---|---|
| The control area (non-LAD LV region) was determined by the area of distribution of the fluorescent microspheres. The normal and collateral-dependent zones were divided with a blade, and for each the total weight was measured. Collateral flow was calculated as a ratio between activity (dpm/g) of the tissue samples from the LAD-dependent and normal zones. | The control area (non-LAD LV region) was determined by the area distribution of the fluorescent microspheres. The normal and collateral-dependent zones were divided with a blade, and each total weight was measured. CBF (mL · min-1 · g-1) in each area was calculated from the following formula: CBF=[(radioactive counts in myocardial specimen)x(blood withdrawal rate)/(radioactive count in blood)]/(weight of myocardial specimen). |
The copied text begins on the previous page: Arc/Fragment_042_16 |
|
| [35.] Arc/Fragment 044 01 - Diskussion Bearbeitet: 26. February 2014, 23:34 Hindemith Erstellt: 26. February 2014, 10:20 (Hindemith) | Arc, Becker et al 1999, Fragment, Gesichtet, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 44, Zeilen: 1-6 |
Quelle: Becker et al 1999 Seite(n): H2241, Zeilen: l.col: 21-31 |
|---|---|
| [Intracellular oxidant stress can be monitored by measuring] changes in fluorescence resulting from intracellular probe oxidation. DHE enters the cell and is oxidized by ROS, particularly superoxide, to yield fluorescent ethidium. Ethidium binds to DNA (Eth-DNA), further amplifying its fluorescence. Eth-DNA fluorescence is generally stable but can be decreased by hydroxyl radical attack. Thus increases in DHE oxidation to Eth-DNA (i.e., increases in Eth-DNA fluorescence) are suggestive of superoxide generation. | Intracellular oxidant stress was monitored by measuring changes in fluorescence resulting from intracellular probe oxidation. Dihydroethidine (DHE, 1–10 μmol/l; Molecular Probes) enters the cell and is oxidized by ROS, particularly superoxide, to yield fluorescent ethidium. Ethidium binds to DNA (Eth-DNA), further amplifying its fluorescence (5). Eth-DNA fluorescence is generally stable but can be decreased by hydroxyl radical attack (27). Thus increases in DHE oxidation to Eth-DNA (i.e., increases in Eth-DNA fluorescence) are suggestive of superoxide generation.
5. Carter,W. O., P. K. Narayanan, and J. P. Robinson. Intracellular hydrogen peroxide and superoxide anion detection in endothelial cells. J. Leukoc. Biol. 55: 253–258, 1994. 27. Prutz, W. A. Inhibition of DNA-ethidium bromide intercalation due to free radical attack upon DNA. Radiat. Environ. Biophys. 23: 1–18, 1984. |
The source is not mentioned. |
|
| [36.] Arc/Fragment 044 15 - Diskussion Bearbeitet: 26. February 2014, 10:54 Hindemith Erstellt: 24. February 2014, 23:26 (Graf Isolan) | Arc, Fragment, Gesichtet, Rocic et al 2007, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 44, Zeilen: 15-25 |
Quelle: Rocic et al 2007 Seite(n): H2730, Zeilen: left col. 33-47 |
|---|---|
| For the in vitro studies in freshly isolated cardiomyocytes (Cardiomyocyte Isolation for details), G-CSF was administered for 10 min (total treatment time 30 min), and DHE was added during the last 20 min of treatment with G-CSF. Cells were then immediately observed under a fluorescent microscope. For in vivo studies, DHE was injected into the LV (60 μg/kg, n=3/group) for 20 min before two consecutive periods of ischemia-reperfusion (40s occlusion followed by 20min reperfusion and another 40s occlusion and 20min reperfusion). Animals were then sacrificed. The heart was removed, frozen in optimum cutting temperature compound on dry ice, and stored at -80°C until sectioning. Sections (5 μm) were made in a cryomicrotome and were mounted on glass slides. DHE fluorescence was detected with excitation/emission at 518/605 nm. All images were analyzed at the same microscope settings, and fluorescence intensity was obtained by Metamorph Software on three hearts (10 sections per heart). | For HCAEC cell culture, DHE was administered during the last 20 min of treatment. Cells were then observed immediately under a fluorescent microscope. For in vivo studies, DHE or DCF were injected into the LV (60 μg/kg) for 20 min before two consecutive periods of ischemia-reperfusion (40 s occlusion followed by 20 min reperfusion and another 40 s occlusion and 20 min reperfusion). Animals were then killed. The heart was removed, frozen in optimum cutting temperature compound on dry ice, and stored at -70°C until sectioning. Sections (5 μm) were made in a cryomicrotome and were mounted on glass slides. DHE or DCF fluorescence was detected with excitation/emission at 518/605 nm (for DHE) or 480/515 nm (for DCF). All images were analyzed at the same microscope settings, and relative fluorescence readings were obtained by Metamorph Software on three hearts (5 consecutive sections per heart). |
Though taken verbatim nothing has been marked as a citation and the source is not given. |
|
| [37.] Arc/Fragment 046 25 - Diskussion Bearbeitet: 26. February 2014, 20:25 Hindemith Erstellt: 26. February 2014, 20:05 (Graf Isolan) | Arc, Fragment, Gesichtet, Rocic et al 2007, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 46, Zeilen: 25-27 |
Quelle: Rocic et al 2007 Seite(n): H2730, Zeilen: right col. 3-8 |
|---|---|
| The extent of tube formation was quantified after 24h by superimposing a grid (25mm2/cube) on microscopic images, and the number of squares containing tubes were counted and [averaged from five randomly selected fields for each well to obtain the percentage of total field that contained tubes.] | The extent of tube formation was quantified after two days by using Scion Image software. An electronic grid was superimposed on microscopic images, and the number of squares containing tubes were counted and averaged from five randomly selected fields for each well to obtain the percentage of total field that contained tubes. |
Nearly identical, but not marked as a citation. |
|
| [38.] Arc/Fragment 047 01 - Diskussion Bearbeitet: 26. February 2014, 22:57 Schumann Erstellt: 26. February 2014, 20:13 (Graf Isolan) | Arc, Fragment, Gesichtet, Rocic et al 2007, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 47, Zeilen: 1-5 (complete) |
Quelle: Rocic et al 2007 Seite(n): H2730, Zeilen: right col. 3-8, 22-26 |
|---|---|
| [The extent of tube formation was quantified after 24h by superimposing a grid (25mm2/cube) on microscopic images, and the number of squares containing tubes were counted and] averaged from five randomly selected fields for each well to obtain the percentage of total field that contained tubes. Treatments were conducted in n=4.
3.9 Data analysis ANOVA followed by t-tests was used for statistical analysis. A probability value of P<0.05 was used to determine statistical significance. |
The extent of tube formation was quantified after two days by using Scion Image software. An electronic grid was superimposed on microscopic images, and the number of squares containing tubes were counted and averaged from five randomly selected fields for each well to obtain the percentage of total field that contained tubes.
[...] All experiments were performed in triplicate (n=3 animals/group). Data analysis. ANOVA followed by t-tests with Bonferroni inequalities was used for statistical analysis. A probability value of P<0.05 was used to determine statistical significance. |
Nearly identical, but not marked as a citation. |
|
| [39.] Arc/Fragment 049 02 - Diskussion Bearbeitet: 26. February 2014, 23:07 Guckar Erstellt: 23. February 2014, 13:45 (Hindemith) | Arc, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Toyota et al 2005, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 49, Zeilen: 2-11 |
Quelle: Toyota et al 2005 Seite(n): 2109, 2110, Zeilen: 2109: r.col: last two lines; 2110: l.col: 1ff |
|---|---|
| Evaluation of collateral growth was performed after completion of the protocol by a simple functional test: total coronary occlusion. The rationale for this procedure was that if collaterals were developed, then occlusion would not induce functional disturbances. Alternatively, if collaterals were not mature, then occlusion would cause hemodynamic disturbances. After the second measurement of coronary blood flow, the coronary occlusion was maintained, systemic hemodynamics were measured and also the number of arrhythmias. In animals without collaterals, coronary occlusion caused deterioration of systemic hemodynamics and arrhythmias, including premature ventricular contractions, ventricular tachycardia, and ventricular fibrillation; in animals with well developed collaterals, no such adverse effects were noted. | We evaluated collateral growth after completion of the protocol by a simple functional test: total coronary occlusion. The rationale for this
[page 2110] procedure was that if collaterals were developed, then occlusion would not induce functional disturbances. Alternatively, if collaterals were not mature, then occlusion would cause hemodynamic disturbances. After the second measurement of coronary blood flow, we maintained the coronary occlusion (n=9 in control group, n=6 in anti-VEGF group) and measured systemic hemodynamics and the number of arrhythmias. In animals without collaterals, coronary occlusion caused deterioration of systemic hemodynamics and arrhythmias, including premature ventricular contractions, ventricular tachycardia, and ventricular fibrillation; in animals with well-developed collaterals, no such adverse effects were noted. |
The source is not mentioned here. |
|
| [40.] Arc/Fragment 049 14 - Diskussion Bearbeitet: 26. February 2014, 23:07 Guckar Erstellt: 23. February 2014, 14:02 (Hindemith) | Arc, Fragment, Gesichtet, SMWFragment, Schutzlevel sysop, Toyota et al 2005, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 49, Zeilen: 14-19 |
Quelle: Toyota et al 2005 Seite(n): 2109, Zeilen: r.col: 36-42 |
|---|---|
| After the completion of treatments, the heart was removed, and the left ventricle (LV) was sliced along the short axis and observed with a dissecting microscope under fluorescent light. The collateral-dependent area (LAD region, ischemic zone) was distinguished as the area without fluorescent microspheres. The control area (non-LAD LV region, normal zone) was determined by the area of distribution of the fluorescent microspheres (Fig. 13). | The heart was excised and fixed in 4% paraformaldehyde solution overnight. The fixed LV was sliced along the short axis and observed with a dissecting microscope and fluorescent light source (LT-9800, Lightools Research). The collateral-dependent area (LAD region) was distinguished as the area without fluorescent microspheres. The control area (non-LAD LV region) was determined by the area distribution of the fluorescent microspheres. |
The source is not given. To be continued on the next page: Arc/Fragment_050_01 Note that a similar text can also be found further up in the thesis, c.f. Arc/Fragment_042_16, Arc/Fragment_043_01 |
|
| [41.] Arc/Fragment 050 01 - Diskussion Bearbeitet: 26. February 2014, 23:07 Guckar Erstellt: 23. February 2014, 14:06 (Hindemith) | Arc, Fragment, Gesichtet, KomplettPlagiat, SMWFragment, Schutzlevel sysop, Toyota et al 2005 |
|
|
|
| Untersuchte Arbeit: Seite: 50, Zeilen: 1-2 |
Quelle: Toyota et al 2005 Seite(n): 2109, Zeilen: r.col: 42-44 |
|---|---|
| The normal and collateral-dependent zones were divided with a blade, and each total weight was measured. | The normal and collateral-dependent zones were divided with a blade, and each total weight was measured. |
The copied text starts on the previous page: Arc/Fragment_049_14 |
|