Diese Zusammenstellung basiert auf Befunden einer laufenden Plagiatsanalyse (Stand: 2014-05-02) – es handelt sich insofern nicht um einen abschließenden Bericht. Zur weiteren Meinungsbildung wird daher empfohlen, den jeweiligen Stand der Analyse auf der Seite http://de.vroniplag.wikia.com/wiki/Aeh zum Vergleich heranzuziehen.
A critical discussion of the thesis by Aziza El Harrak: Pharmacological investigation of spreading depression propagation in rat neocortical tissues
Submitted to the department of medicine of the Wilhelms-Universität Münster, in fulfillment of the requirements for the degree of doctor medicinae dentium. 1st examiner: PD Dr. med. Christoph Greiner 2nd examiner: Prof. Dr. med. Ali Gorji. Oral examination: 20. November 2009. Publication: Münster, 6. January 2010.
→ Nachweis: Deutsche Nationalbibliothek "Ursprünglich als Dissertation veröffentlicht, Doktorgrad wurde am 28.04.2016 entzogen."
→ Nachweis: UB Münster "Entzug des Doktorgrades im Jahr 2016."
→ Verwaltungsgericht Münster weist am 13. März 2017 die Klage ab (Az 10 K 1792/16)
→ Oberverwaltungsgericht Nordrhein-Westfalen weist am 4.1.2018 die Berufung zurück (14 A 610/17)
Der Barcode drückt den Anteil der Seiten aus, die Fremdtextübernahmen enthalten, nicht den Fremdtextanteil am Fließtext. Je nach Menge des übernommenen Textes werden drei Farben verwendet:
- schwarz: bis zu 50 % Fremdtextanteil auf der Seite
- dunkelrot: zwischen 50 % und 75 % Fremdtextanteil auf der Seite
- hellrot: über 75 % Fremdtextanteil auf der Seite
Weiße Seiten wurden entweder noch nicht untersucht oder es wurde nichts gefunden. Blaue Seiten umfassen Titelblatt, Inhaltsverzeichnis, Literaturverzeichnis, Vakatseiten und evtl. Anhänge, die in die Berechnung nicht einbezogen werden.
Der Barcode stellt den momentanen Bearbeitungsstand dar. Er gibt nicht das endgültige Ergebnis der Untersuchung wieder, da Untersuchungen im VroniPlag Wiki stets für jeden zur Bearbeitung offen bleiben, und somit kein Endergebnis existiert.
13 Seiten mit Plagiatstext
Seiten mit weniger als 50% Plagiatstext
Seiten mit 50%-75% Plagiatstext
Seiten mit mehr als 75% Plagiatstext
8 Seiten: 010 009 019 007 008 018 017 006
Findings
- Problematic text parallels can be found in the following chapters (state of analysis: 02.05.2014):
- Material and methods
- Results
- Control slices (p. 15): page 15
Prominent sources
- Gorji (2001): Roughly two pages of the discussion section have been taken from a review paper authored by the second examiner of the thesis Prof. Dr. Gorji. This includes 18 references to the literature.
- Sheikh (2009): Most of the remaining part of the discussion section seems to be taken from a dissertation that has been submitted a bit less than a year earlier in Münster. The first examiner of that dissertation is the second examiner of Aeh.
- Herreras et al. (1994): More than a page of the introduction section has been taken from a paper that was about 15 years old at the time of writing of the thesis. As a consequence the references to the literature taken from this source appear quite dated.
Prominent Fragments
- Fragment 018 01: An entire page in the discussion section has been copied without attribution.
- Fragment 009 14: A clear indication that text has been taken via copy & paste.
Other observations
- The second examiner of the thesis Prof. Dr. Gorji was the supervisor of the dissertation Sheikh (2009) as well as author or co-author of the publications Gorji (2001), Gorji et al. (2003) and Wernsmann et al. (2006), so it would have been possible for him to spot the quite substantial text parallels between the dissertation Aeh (2009) and those sources.
- The regulations for the submission of dissertations in effect at the time demand a declaration accompanying the thesis that "die Doktorandin/ der Doktorand sie nur unter Benutzung der im Literaturverzeichnis angegebenen Quellen angefertigt hat und sonst kein anderes gedrucktes oder ungedrucktes Material verwendet wurde" (§2 (1) 4.)
- It seems surprising that a doctoral degree in dentistry ("doctor medicinae dentium" according to cover page) is granted for a thesis studying neural mechanisms in the rat brain.
- The dissertation Haarmann (2009) has been prepared at the same time as Aeh (the date of the oral examination is the same for both theses: 20.11.2009). It contains substantial text parallels with Aeh that, however, have not been documented here, because it is not clear who has copied from whom. The second paragraph on page 6 (lines 17-25) for instance has a parallel in Haarmann (2009) (page 5: 16ff).
Statistics
- Currently there are 21 reviewed fragments documented that are considered to be violations of citation rules. For 20 of them there is no reference given to the source used („Verschleierungen“ and „Komplettplagiate“). For 1 fragments the source is given, but the extent of the used text is not made clear („Bauernopfer“).
- The publication has 15 pages that have been analyzed. On a total of 13 of these pages violations of citation rules have been documented. This represents a percentage of 86.7%. The 15 analyzed pages break down with respect to the amount of text parallels encountered as follows:
- From these statistics an extrapolation of the amount of text of the publication under investigation that has been documented as problematic can be estimated (conservatively) as about 55% of the main part of the publication.
- In all, text was taken from 7 sources.
Illustration
The following chart illustrates the extent and the distribution of the text parallels found. The colours show the type of violation of citation rules diagnosed:
- grau="Komplettplagiat" (copy & paste): the source of the text parallel is not given, the copy is verbatim.
- rot="Verschleierung" (disguised copy): the source of the text parallel is not given, the copied text will be somewhat modified.
- gelb="Bauernopfer" (pawn sacrifice): the source of the text parallel is mentioned, but the extent and/or the closeness of the copy to the source is not made clear by the reference.

(state of analysis: 02.05.2014)
Definition von Plagiatkategorien
Die hier verwendeten Plagiatkategorien basieren auf den Ausarbeitungen von Wohnsdorf / Weber-Wulff: Strategien der Plagiatsbekämpfung, 2006. Eine vollständige Beschreibung der Kategorien findet sich im VroniPlag-Wiki. Die Plagiatkategorien sind im Einzelnen:
Übersetzungsplagiat
Ein Übersetzungsplagiat entsteht durch wörtliche Übersetzung aus einem fremdsprachlichen Text. Natürlich lässt hier die Qualität der Übersetzung einen mehr oder weniger großen Interpretationsspielraum. Fremdsprachen lassen sich zudem höchst selten mit mathematischer Präzision übersetzen, so dass jede Übersetzung eine eigene Interpretation darstellt. Zur Abgrenzung zwischen Paraphrase und Kopie bei Übersetzungen gibt es ein Diskussionsforum.
Komplettplagiat
Text, der wörtlich aus einer Quelle ohne Quellenangabe übernommen wurde.
Verschleierung
Text, der erkennbar aus fremder Quelle stammt, jedoch umformuliert und weder als Paraphrase noch als Zitat gekennzeichnet wurde.
Bauernopfer
Text, dessen Quelle ausgewiesen ist, der jedoch ohne Kenntlichmachung einer wörtlichen oder sinngemäßen Übernahme kopiert wurde.
Quellen nach Fragmentart
Die folgende Tabelle schlüsselt alle gesichteten Fragmente zeilenweise nach Quellen und spaltenweise nach Plagiatskategorien auf.
- ÜP = Übersetzungsplagiat,
- KP = Komplettplagiat,
- VS = Verschleierung,
- BO = Bauernopfer,
- KW = Keine Wertung,
- KeinP = Kein Plagiat.
| Quelle |
Jahr | ÜP |
KP |
VS |
BO |
KW |
KeinP |
∑ |
ZuSichten |
Unfertig |
|---|---|---|---|---|---|---|---|---|---|---|
| Gorji | 2001 | 0 | 3 | 2 | 1 | 0 | 0 | 6 | 0 | 0 |
| Gorji et al | 2003 | 0 | 2 | 1 | 0 | 0 | 0 | 3 | 0 | 0 |
| Herreras et al | 1994 | 0 | 1 | 1 | 0 | 0 | 0 | 2 | 0 | 0 |
| Sheikh | 2009 | 0 | 4 | 2 | 0 | 0 | 0 | 6 | 0 | 0 |
| Smith et al | 2006 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Somjen | 2001 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Wernsmann et al | 2006 | 0 | 1 | 1 | 0 | 0 | 0 | 2 | 0 | 0 |
| ∑ | - | 0 | 11 | 9 | 1 | 0 | 0 | 21 | 0 | 0 |
Fragmentübersicht
21 gesichtete, geschützte Fragmente
| Fragment | SeiteArbeit | ZeileArbeit | Quelle | SeiteQuelle | ZeileQuelle | Typus |
|---|---|---|---|---|---|---|
| Aeh/Fragment 006 05 | 6 | 5-10, 12-16 | Smith et al 2006 | 459 | l.col: 2ff | Verschleierung |
| Aeh/Fragment 006 17 | 6 | 17-25 | Somjen 2001 | 1066, 1067 | 1066: r.col: last 3 lines; 1067: l.col: 1ff | Verschleierung |
| Aeh/Fragment 007 02 | 7 | 2-18 | Gorji 2001 | 34 | l.col: 7ff | BauernOpfer |
| Aeh/Fragment 007 19 | 7 | 19-25 | Sheikh 2009 | 7 | 16-22 | Verschleierung |
| Aeh/Fragment 008 01 | 8 | 1-2 | Sheikh 2009 | 8, 9 | 8: 1-2; 9: figure | KomplettPlagiat |
| Aeh/Fragment 008 03 | 8 | 3-5 | Gorji 2001 | 42 | r.col: 46ff | KomplettPlagiat |
| Aeh/Fragment 009 01 | 9 | 1-12 | Gorji 2001 | 42, 43 | 42: r.col: 47ff - 43: l.col: 1ff | KomplettPlagiat |
| Aeh/Fragment 009 14 | 9 | 14-25 | Herreras et al 1994 | 7087 | r.col: 8ff | Verschleierung |
| Aeh/Fragment 010 01 | 10 | 1-21 | Herreras et al 1994 | 7087, 7097 | 7087: r.col: 27ff; 7097: l.col: 2ff | KomplettPlagiat |
| Aeh/Fragment 010 22 | 10 | 22-25 | Gorji et al 2003 | 113 | l.col: 20ff | KomplettPlagiat |
| Aeh/Fragment 011 01 | 11 | 1-6 | Gorji et al 2003 | 113 | l.col: 25ff | KomplettPlagiat |
| Aeh/Fragment 012 08 | 12 | 8-15 | Wernsmann et al 2006 | 1104 | l.col: 4-12 | KomplettPlagiat |
| Aeh/Fragment 012 16 | 12 | 16-22 | Gorji et al 2003 | 114 | l.col: 49ff | Verschleierung |
| Aeh/Fragment 013 01 | 13 | 1-7 | Wernsmann et al 2006 | 1104 | l.col: 22-31 | Verschleierung |
| Aeh/Fragment 015 03 | 15 | 3-6, 8-11 | Sheikh 2009 | 14 | 10-17 | Verschleierung |
| Aeh/Fragment 017 06 | 17 | 6-16 | Sheikh 2009 | 22 | 7-19 | KomplettPlagiat |
| Aeh/Fragment 017 17 | 17 | 17-23 | Gorji 2001 | 38 | l.col: 20ff | Verschleierung |
| Aeh/Fragment 018 01 | 18 | 1ff (complete) | Gorji 2001 | 38 | l.col: 30ff | Verschleierung |
| Aeh/Fragment 019 01 | 19 | 1-15 | Gorji 2001 | 38 | r.col: 14ff | KomplettPlagiat |
| Aeh/Fragment 019 19 | 19 | 19-25 | Sheikh 2009 | 23, 24 | 23: 24ff - 24: 1-5 | KomplettPlagiat |
| Aeh/Fragment 020 01 | 20 | 1-9 | Sheikh 2009 | 24 | 5-13 | KomplettPlagiat |
Textfragmente
Anmerkung zur Farbhinterlegung
Die Farbhinterlegung dient ausschließlich der leichteren Orientierung des Lesers im Text. Das Vorliegen einer wörtlichen, abgewandelten oder sinngemäßen Übernahme erschließt sich durch den Text.
Hinweis zur Zeilenzählung
Bei der Angabe einer Fundstelle wird alles, was Text enthält (außer Kopfzeile mit Seitenzahl), als Zeile gezählt, auch Überschriften. In der Regel werden aber Abbildungen, Tabellen, etc. inklusive deren Titel nicht mitgezählt. Die Zeilen der Fußnoten werden allerdings beginnend mit 101 durchnummeriert, z. B. 101 für die erste Fußnote der Seite.
21 gesichtete, geschützte Fragmente
| [1.] Aeh/Fragment 006 05 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 6, Zeilen: 5-10, 12-16 |
Quelle: Smith et al 2006 Seite(n): 459, Zeilen: l.col: 2ff |
|---|---|
| It was first induced by applying a brief tetanus of faradic stimulation to the rabbit cortex (Leao, 1944; Bures et al., 1974). Mechanical stimulation, for example, by stroking of the cortical surface with a blunt instrument, a falling weight or even lightly tapping the cortex also initiates SD (Lea˜o [sic], 1944). More recent studies have achieved more reliable and reproducible induction of SD by rapidly inserting and retracting hypodermic steel needles (Lambert et al., 1999; Ebersberger et al., 2001). [...] This model has been shown to be the most reliable stimulus leading to reproducible events on earlier occasions in both non-imaging and imaging studies (Bradley et al., 2001). In any case, changes in extracellular K+ concentration themselves might be involved in such pathophysiological processes in human brain tissue (Mayevsky et al., 1996; Nicholson and Sykova, 1998). SD appears to affect both the neuronal and the glial cells. | The CSD phenomenon is exclusive to the central nervous system (CNS) and appears to involve both the neuronal and the glial cell populations. [...] CSD was first induced by applying a brief tetanus of faradic stimulation to the rabbit cortex (Leão, 1944; Bureš, Buresová & Krivánek, 1974). [...]
Mechanical stimulation, for example, by stroking of the cortical surface with a blunt instrument, a falling weight or even lightly tapping the cortex also evokes CSD (Leão, 1944; Zachar & Zacharová, 1963). More recent studies have achieved more reliable and reproducible induction of CSD by rapidly inserting and retracting hypodermic steel needles (Kaube & Goadsby, 1994; Lambert et al., 1999; Ebersberger et al., 2001), [...] Of available methods KCl has thus proven to be the most reliable stimulus leading to reproducible events on earlier occasions in both non-imaging (Bureš et al., 1974, 1984; Lehmankühler & Richter, 1993; Smith et al., 1998, 2000; Read et al., 1999; Martins-Ferreira et al., 2000; Kuge et al., 2000) and imaging studies (Gardner-Medwin et al., 1994; Latour et al., 1994;Hasegawa et al., 1995; de Crespigny et al., 1996, 1998; James et al., 1999; Bockhorst et al., 2000; Kuge et al., 2000; Bradley et al., 2001, 2002). In any case, changes in extracellular K+ concentration ([K+])o themselves might be involved in such pathophysiological processes in human brain tissue (Mayevsky et al., 1996; Nicholson & Sykova´, 1998). |
The source is not mentioned. |
| [2.] Aeh/Fragment 006 17 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 6, Zeilen: 17-25 |
Quelle: Somjen 2001 Seite(n): 1066, 1067, Zeilen: 1066: r.col: last 3 lines; 1067: l.col: 1ff |
|---|---|
| The first paper on SD, titled "Spreading depression of activity in the cerebral cortex" appeared in 1944, written by a Brazilian neuroscientist, Aristides Leão, working at the Harvard laboratory. Leão wanted to study the electrocorticogram (ECoG) of experimental epilepsy in anesthetized rabbits, but he was distracted from his original goal by an unexpected flattening of the ongoing normal bioelectrical activity that took the place of the anticipated epileptiform field potentials. The silencing of the ECoG trace crept slowly over the cortex, from one recording electrode pair resting on the cortical surface to the one beside it. According to Leão, SD and propagating focal seizures were related phenomena, generated by the same cellular elements, an inference later supported by others. | The first, seminal paper on SD, titled “Spreading depression of activity in the cerebral cortex” (213) appeared in 1944, written by a young and unknown Brazilian inves-
[page 1067] tigator, Aristides Leão, working at the Harvard laboratory of R. S. Morison. Leão wanted to study the cortical electrogram (ECoG) of experimental epilepsy in anesthetized rabbits, but he was distracted from his original goal by an unexpected silencing of the ongoing normal electrical activity that took the place of the anticipated seizure (Fig. 1). The flattening of the ECoG trace crept slowly over the cortex, from one recording electrode pair resting on the cortical surface to the one beside it. According to Leão, SD and propagating focal seizures were related phenomena, generated by the same cellular elements (213), an inference later supported by others (e.g., Ref. 428). [...] |
The source is indicated in the bibliographry, but no reference is made in the present context. |
| [3.] Aeh/Fragment 007 02 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 7, Zeilen: 2-18 |
Quelle: Gorji 2001 Seite(n): 34, Zeilen: l.col: 7ff |
|---|---|
| SD appears first at the stimulated site and spreads out in all directions at the velocity of 2–3 mm/min, so that increasingly distant areas undergo successively a similar temporary depression. A crucial manifestation of SD is a propagating negative potential with amplitude of 10–30 mV and duration of more than 30 sec, which may be preceded or succeeded by a positive fluctuation of variable amplitude and duration (Figure 1). Underlying this cellular depolarisation is a dramatic change in the distribution of micromilieu ions between extra- and intracellular compartments. Potassium and proton release from the cells, while sodium, calcium and chloride enter together with water causing cells to swell and the volume of the extracellular compartment to be decreased. SD is accompanied by an increase of glucose utilization and O2 consumption. Recovery of SD depends on energy metabolism.
SD has been studied in vivo and in vitro in brain slices and in retinal preparations under different experimental conditions. It has been also observed in human neocortical tissue in vitro and in human hippocampus as well as striatum and neocortex in vivo. SD can be regularly initiated if the tissue susceptibility is artificially raised. Hypoxia as well as hypoglycemia and changing the extracellular ionic concentrations by administration of solutions with increased K+, decreased NaCl or with the Cl− of the latter replaced by certain other anions lower the threshold (Gorji, 2001). |
It appears first at the stimulated site and spreads out in all directions at the velocity of 2–3 mm/min, so that increasingly distant areas undergo successively a similar temporary depression [243]. A necessary manifestation of SD is a propagating extracellular negative potential with an amplitude of 10-30 mV and a duration of more than 0.5-1 min, which may be preceded or succeeded by a positive deflection of variable amplitude and duration. Underlying this neuro-glial depolarization is a dramatic change in the distribution of ions between extra- and intracellular spaces. K+ and H+ release from the cells, while Na+, Ca2+ and Cl- enter together with water [152,166,222] causing cells to swell and the volume of the extracellular compartment to be reduced. SD is accompanied by an increase of glucose utilization and O2 consumption [47,283]. Recovery of SD depends on energy metabolism [47].
This phenomenon has been studied in vivo in several animal species and in vitro in brain slices and in retinal preparations under various experimental conditions [47]. It has been also observed in human neocortical tissue in vitro [16,17,149] and in human hippocampus as well as striatum [408] and neocortex [272] in vivo. [...] SD can be regularly initiated if the tissue susceptibility is artificially raised. Hypoglycemia and hypoxia as well as changing the extracellular ionic micromilieu by applying solutions with increased K+, decreased NaCl or with the Cl- of the latter replaced by certain other anions lower the threshold. |
The source is mentioned at the end, but it is not clear that the preceding two(!) paragraphs are taken from it. |
| [4.] Aeh/Fragment 007 19 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 7, Zeilen: 19-25 |
Quelle: Sheikh 2009 Seite(n): 7, Zeilen: 16-22 |
|---|---|
| There is sufficient evidence to admit the SD plays an important role in different neurological disorders (Gorji, 2001; Somjen, 2001). Subdural recordings in patients demonstrated that SD is critically involved in various disorders associated with acute neuronal injury including traumatic and spontaneous intracerebral haemorrhage (Strong et al., 2002; Fabricius et al., 2008) as well as subarachnoid haemorrhage and ischaemic stroke and contribute to tissue damage. Furthermore, propagation of a SD-like phenomenon in human neocortical tissues has been shown to generate aura symptoms in migrainous patients (Hadjikhani et al. 2001). | There is sufficient evidence to admit the SD plays an important role in different neurological disorders (Gorji, 2000). Subdural recordings in patients demonstrated that SD is critically involved in various disorders associated with acute neuronal injury including traumatic and spontaneous intracerebral haemorrhage (Strong et al., 2002; Fabricius et al., 2006) as well as subarachnoid haemorrhage and ischaemic stroke (Dreier et al., 2006) and contribute to tissue damage. Furthermore, propagation of a SD-like phenomenon in human neocortical tissues has been shown to generate aura symptoms in migrainous patients (Hadjikhani et al. 2001). |
The source is not mentioned. Some of the literature given is different. |
| [5.] Aeh/Fragment 008 01 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 8, Zeilen: 1-2 |
Quelle: Sheikh 2009 Seite(n): 8, 9, Zeilen: 8: 1-2; 9: figure |
|---|---|
| Although interrelation of SD and epilepsy has been considered for a long time, the possible pathophysiological role of SD in epilepsy needs to be elucidated.
Figure 1. Vertical propagation of a negative DC-potential wave after injection of KCl in a neocortical slice. Injection of KCl solution (3 M) via a microelectrode elicited spreading depression-like fluctuation during superfusion with artificial cerebrospinal fluid. Injecting and recording electrodes arranged as shown. Voltage variations were recorded simultaneously by four electrodes (DC1–DC4) which set apart by 1 mm (Adopted from Gorji et al., 2001). |
Although interrelation of SD and epilepsy has been considered for a long time, the possible pathophysiological role of SD in epilepsy needs to be elucidated.
Fig. 1. Propagation of a negative DC-potential wave after injection of KCl in a neocortical slice. Injection of KCl solution (3 M) via a microelectrode elicited spreading depression-like fluctuation during superfusion with artificial cerebrospinal fluid. Injecting and recording electrodes arranged as shown. Voltage variations were recorded simultaneously by four electrodes (DC1–DC4) which set apart by 1 mm. |
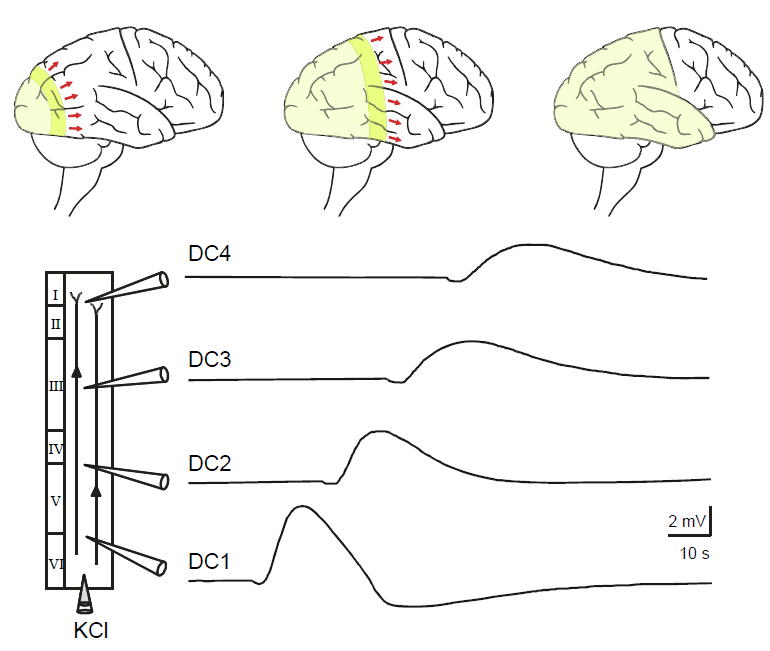
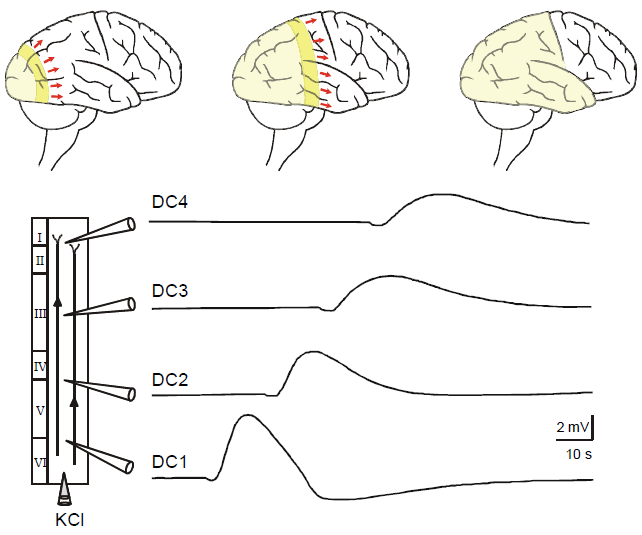
The copied text starts on the previous page: Aeh/Fragment_007_19. Note that in Gorji et al., (2001) there is a very similar but slightly different figure with a slightly different figure caption. |
| [6.] Aeh/Fragment 008 03 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 8, Zeilen: 3-5 |
Quelle: Gorji 2001 Seite(n): 42, Zeilen: r.col: 46ff |
|---|---|
| SD is also a well-known phenomenon in experimental epilepsy. SD has been observed in a variety of in vitro and in vivo epilepsy models in different animal species. Reduction of extracellular Mg2+ concentrations, activation of NMDA receptors, blocking of K+ channels, [e.g., by 4-aminopyridine, increased extracellular K+, blocking of Na+–K+ ATPase, e.g., by ouabain, blocking of Ca2+ channels, e.g., by NiCl2, blocking of GABA receptors, e.g., by picrotoxin, are the common pathways for eliciting epileptiform burst discharges and SD in experimental models.] | SD is a well-known phenomenon in experimental epilepsy. SD has been observed in a variety of in vitro and in vivo epilepsy models in different animals [sic] species. Reduction of extracellular Mg2+ concentrations, activation of NMDA receptors, blocking of K+ channels, e.g., by 4-aminopyridine, increased extracellular K+ , blocking of Na+-K+ ATPase, e.g., by ouabain, blocking of Ca2+ channels, e.g., by NiCl2, blocking of GABA receptors, e.g., by picrotoxin, are the common pathways for eliciting epileptiform burst discharges and SD in experimental models [22,149,150,243,337,344,433]. |
The source is not mentioned. |
| [7.] Aeh/Fragment 009 01 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 9, Zeilen: 1-12 |
Quelle: Gorji 2001 Seite(n): 42, 43, Zeilen: 42: r.col: 47ff - 43: l.col: 1ff |
|---|---|
| [SD has been observed in a variety of in vitro and in vivo epilepsy models in different animal species. Reduction of extracellular Mg2+ concentrations, activation of NMDA receptors, blocking of K+ channels,] e.g., by 4-aminopyridine, increased extracellular K+, blocking of Na+–K+ ATPase, e.g., by ouabain, blocking of Ca2+ channels, e.g., by NiCl2, blocking of GABA receptors, e.g., by picrotoxin, are the common pathways for eliciting epileptiform burst discharges and SD in experimental models. By all aforementioned mechanisms SD appears spontaneously between epileptiform ictal events. SD can be elicited in susceptible area by a single discharge of an epileptic focus (spike triggered SD). Epileptiform field potentials usually suppress during SD occurrence and reappear in few minutes. CSD penetration into the epileptic foci was established in different model of epilepsy. However, it should be noted that SD does not enter electrically or pharmacologically elicited foci of epileptic activity with high rates of interictal discharges which resulted in anomalous SD propagation. This abnormal SD conduction may account for periodic changes of ictal and interictal activity found in some types of focal epilepsy. | SD has been observed in a variety of in vitro and in vivo epilepsy models in different animals [sic] species. Reduction of extracellular Mg2+ concentrations, activation of NMDA receptors, blocking of K+ channels, e.g., by 4-aminopyridine, increased extracellular K+ , blocking of Na+-K+ ATPase, e.g., by ouabain, blocking of Ca2+ channels, e.g., by NiCl2, blocking of GABA receptors, e.g., by picrotoxin, are the common pathways for eliciting epileptiform burst discharges and SD in experimental models [22,149,150,243,337,344,433]. By all aforemen-
[page 43] tioned mechanisms SD appears spontaneously between epileptiform ictal events. SD can be elicited in susceptible area by a single discharge of an epileptic focus (spike triggered SD). Epileptiform field potentials usually suppress during SD occurrence and reappear in few minutes [150,217]. CSD penetration into epileptic foci established in different model of epilepsy. However, it should be noted that SD does not enter electrically or pharmacologically elicited foci of epileptic activity with high rates of interictal discharges which resulted in anomalous SD propagation. This abnormal SD conduction may account for periodic changes of ictal and interictal activity found in some types of focal epilepsy [50,215,217]. [...] |
The source is not mentioned. |
| [8.] Aeh/Fragment 009 14 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 9, Zeilen: 14-25 |
Quelle: Herreras et al 1994 Seite(n): 7087, Zeilen: r.col: 8ff |
|---|---|
| Widely accepted hypotheses hold that the primary event responsible for both the initiation and the propagation of SD is the release of some substance from neuronal elements to the extracellular compartment, which initially excites and then depresses adjacent neurons. The slowness of diffusion of the mediator would account for the low velocity of SD propagation. Among the substances proposed to mediate SD propagation are potassium (Grafstein, 1963; Bures et al., 1974) and excitatory amino acids (Fabricius et al., 1993). There are, however, observations that are difficult to reconcile with either of these two propositions.
SD had been interpreted as a composite process or a sequence of several linked events. To solve its genesis, a most important question concerns identification of the very first step in the chain reaction. In the extant literature, however, generally more attention has been given to the major depolarization and the attending extracellular potential shift (AI’,) [sic] than to the antecedent events. |
Widely accepted hypotheses hold that the primary event responsible for both the initiation and the propagation of SD is the release of some substance from neuronal elements to the extracellular compartment, which initially excites and then depresses adjacent neurons. The slowness of diffusion of the mediator would account for the low velocity of SD propagation. Among the substances proposed to mediate SD propagation are potassium (Grafstein, 1956; Brinley et al., 1960; BureS and Kiivanek, 1960) and excitatory amino acids (Van Harreveld, 1959; SiesjG and Bengtsson, 1989; Fabricius et al., 1993). There are, however, observations that are difficult to reconcile with either of these two propositions (Lehmenkiihler, 1990; Herreras and Somjen, 1993a,b; see also Discussion).
SD had been interpreted as a composite process or a sequence of several linked events. To solve its genesis, a most important question concerns identification of the very first step in the chain reaction. In the extant literature, however, generally more attention has been given to the major depolarization and the attending extracellular potential shift (ΔV0) than to the antecedent events. |
The source is not mentioned. Note, if one takes the online available version of the source and copies (ΔV0) and then pastes that expression, one obtains: (AI’,) |
| [9.] Aeh/Fragment 010 01 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 10, Zeilen: 1-21 |
Quelle: Herreras et al 1994 Seite(n): 7087, 7097, Zeilen: 7087: r.col: 27ff; 7097: l.col: 2ff |
|---|---|
| [Among antecedents heralding the onset of SD that have been reported, are] a slight increase of extracellular potassium, a small positive shift preceding the fast negative shift of the extracellular potential and several types of fast field activity including a short burst of action potentials or intense synaptic noise (Leao, 1944; Higashida et al., 1974). Even a silence of spontaneous or evoked activity has occasionally been described prior to other signs (Higashida et al., 1974). Not all of these early signs are obligatory prodromals of the large, accelerating, regenerative depolarization that is typical of the process.
Even though the discharge of impulses is not required for the initiation or the propagation of SD, the impulse shower does regularly appear at its beginning. The mechanism that gives rise to the impulse discharge may well have a key role in the evolution of SD. The widely spread synchronization seems best explained by electrical continuity that could be provided by gap junctions. Effective communication by way of quasi-syncytial nets has been demonstrated in other systems, for example in the spread of so-called calcium waves in cell cultures (Cornell-Bell et al., 1990). Patent gap junctions may provide a path not only for electric current and for ions but also for intracellular “second” messengers and other active ingredients in cytosol. SD has frequently been interpreted as a diffusion-reaction process whose velocity of spread is governed by the rate of the reaction, which could involve the release of some substance from cells that then acted on the cell membrane of adjacent cells. As there are reasons for doubting a decisive role of either K or of glutamate, we are proposing an alternative hypothesis, involving the exchange of chemical signals not through the interstitial space but by way of gap junctions. The autocatalytic reaction so initiated would alter the membrane from the inside, instead of acting on receptors on the outside. |
Among antecedents heralding the onset of SD that have been reported, are a slight increase of extracellular potassium ([K+]0), a small positive shift preceding the fast negative shift of the extracellular potential (ΔV0) (Marshall, 1959) and several types of fast field activity including a short burst of action potentials or intense synaptic noise (Leao, 1944; Grafstein 1956; Rosenblueth and Garcia-Ramos, 1966; Ichijo and Ochs, 1970; Higashida et al., 1974; Somjen and Aitken, 1984; Haglund and Schwartzkroin, 1990; Herreras and Somjen, 1993a). Even a silence of spontaneous or evoked activity has occasionally been described prior to other signs (Grafstein, 1956; Morlock et al., 1964; Muiioz, 1970; Higashida et al., 1974). Not all of these early signs are obligatory prodromals of the large, accelerating, regenerative depolarization that is typical of the process.
[page 7097] Even though the discharge of impulses is not required for the initiation or the propagation of SD, the impulse shower does regularly appear at its beginning. The mechanism that gives rise to the impulse discharge may well have a key role in the evolution of SD. The widely spread synchronization seems best explained by electrical continuity that could be provided by gap junctions. Effective communication by way of quasi-syncytial nets has been demonstrated in other systems, for example in the spread of so-called calcium waves in cell cultures (Cornell-Bell et al., 1990; Dani et al., 1992; Finkbeiner, 1992). Patent gap junctions may provide a path not only for electric current and for ions but also for intracellular “second” messengers and other active ingredients in cytosol. SD has frequently been interpreted as a diffusion-reaction process whose velocity of spread is governed by the rate of the reaction, which could involve the release of some substance from cells that then acted on the cell membrane of adjacent cells. As there are reasons for doubting a decisive role of either K or of glutamate, we are proposing an alternative hypothesis, involving the exchange of chemical signals not through the interstitial space but by way of gap junctions. The autocatalytic reaction so initiated would alter the membrane from the inside, instead of acting on receptors on the outside. |
The source is not mentioned. |
| [10.] Aeh/Fragment 010 22 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 10, Zeilen: 22-25 |
Quelle: Gorji et al 2003 Seite(n): 113, Zeilen: l.col: 20ff |
|---|---|
| Cortical structures are organized to process information in a parallel manner via excitatory and inhibitory interactions within and between adjacent cortical modules (Mountcastle, 1997). Throughout the CNS, local circuit inhibition plays an integral role in both neuronal network processing and the regulation of the excitability of projection neurons.
Mountcastle VB. The columnar organization of the neocortex. Brain 1997; 120: 701-722. |
Cortical structures are organized to process information in a parallel manner via excitatory and inhibitory interactions within and between adjacent cortical modules [33]. Throughout the CNS, local circuit inhibition plays an integral role in both neuronal network processing and the regulation of the excitability of projection neurons.
33 Mountcastle VB. The columnar organization of the neocortex. Brain 1997; 120: 701-722 |
The source is not mentioned. |
| [11.] Aeh/Fragment 011 01 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 11, Zeilen: 1-6 |
Quelle: Gorji et al 2003 Seite(n): 113, Zeilen: l.col: 25ff |
|---|---|
| [Inhibitory circuits may] be particularly important to signal processing in cortical networks with pronounced recurrent excitatory interactions (Wong et al., 1984). This inhibition may limit the lateral spread of excitation and facilitate discharge synchronization of projection neurons by inducing a synchronous refractory period. Breakdown in the dynamic balance of inhibitory and excitatory interaction can lead to a functional disconnection (Wong and Prince, 1990) and disrupts the normal spread of lateral excitation (Grunze et al., 1996).
Grunze HC, Rainnie DG, Hasselmo ME, Barkai E, Hearn EF, McCarley RW, Greene RW. NMDA-dependent modulation of CA1 local circuit inhibition. J Neurosci 1996; 16: 2034-2043. Wong BY, Prince DA. The lateral spread of ictal discharges in neocortical brain slices. Epilepsy Res 1990; 7: 29-39. Wong RK, Miles R, Traub RD. Local circuit interactions in synchronization of cortical neurones. J Exp Biol 1984; 112: 169-178. |
Inhibitory circuits may be particularly important to signal processing in cortical networks with pronounced recurrent excitatory interactions [53]. This inhibition may limit the lateral spread of excitation and facilitate discharge synchronization of projection neurons by inducing a synchronous refractory period. Breakdown in the dynamic balance of inhibitory and excitatory interaction can lead to a functional disconnection [52] and disrupts the normal spread of lateral excitation [20].
20 Grunze HC, Rainnie DG, Hasselmo ME, Barkai E, Hearn EF, McCarley RW, Greene RW. NMDA-dependent modulation of CA1 local circuit inhibition. J Neurosci 1996; 16: 2034-2043 52 Wong BY, Prince DA. The lateral spread of ictal discharges in neocortical brain slices. Epilepsy Res 1990; 7: 29-39 53 Wong RK, Miles R, Traub RD. Local circuit interactions in synchronization of cortical neurones. J Exp Biol 1984; 112: 169-178 |
The source is not mentioned. |
| [12.] Aeh/Fragment 012 08 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 12, Zeilen: 8-15 |
Quelle: Wernsmann et al 2006 Seite(n): 1104, Zeilen: l.col: 4-12 |
|---|---|
| Slices were stored at 28 °C in ACSF, which contained (in mm) NaCl, 124; KCl, 4; CaCl2, 1.0; NaH2PO4, 1.24; MgSO4, 1.3; NaHCO3, 26; glucose, 10 (pH 7.4), oxygenated with 95% O2 and 5% CO2 for > 1 h. After 30-min incubation, CaCl2 was elevated to 2.0 mmol/L. Slices were individually transferred to an interphase recording chamber, placed on a transparent membrane, illuminated from below and continuously perfused (1.5–2 mL/min) with carbogenated ACSF at 32 °C. A warmed, humified 95% O2 and 5% CO2 gas mixture was directed over the surface of the slices. | Slices were stored at 28 °C in artificial cerebrospinal fluid (ACSF), which contained (in mM) NaCl, 124; KCl, 4; CaCl2, 1.0; NaH2PO4, 1.24; MgSO4, 1.3; NaHCO3, 26; glucose, 10 (pH 7.4), oxygenated with 95% O2 and 5% CO2 for > 1 h. After 30-min incubation, CaCl2 was elevated to 2.0 mmol/L. Slices were individually transferred to an interphase recording chamber, placed on a transparent membrane, illuminated from below and continuously perfused (1.5–2 mL/min) with carbogenated ACSF at 32 °C. A warmed, humified 95% O2 and 5% CO2 gas mixture was directed over the surface of the slices. |
The source is not given. On the one hand this is the description of an experimental procedure where the variation of wordings is neither easy nor desirable. On the other hand mentioning the study where this procedure has been employed in exactly the same way before would give credit to that study and would aid the interpretation of results. |
| [13.] Aeh/Fragment 012 16 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 12, Zeilen: 16-22 |
Quelle: Gorji et al 2003 Seite(n): 114, Zeilen: l.col: 49ff |
|---|---|
| Electrophysiological recordings
Simultaneous extracellular field potentials were recorded via two glass microelectrodes (150 mmol/L NaCl; 2 - 10 MΩ) positioned at approximately 5-10 mm intervals across the length of the slice in the third cortical layer (figure 2). The reference electrode and the connection to the microelectrode were symmetric Ag-Ag-KCl bridges. Field potentials were traced by an ink writer and recorded by a digital oscilloscope. Inter-electrode distances were measured with a calibrated eyepiece through the microscope. |
Electrophysiological recordings
Simultaneous extracellular field potentials were recorded via four glass microelectrodes (150 mmol/L NaCl; 2 - 10 MΩ) positioned at approximately 2 - 3 mm intervals across the length of the slice in the third cortical layer. [...] The reference electrode and the connection to the microelectrode were symmetric Ag-Ag-KCl bridges. Field potentials were traced by an ink writer and recorded by a digital oscilloscope. Inter-electrode distances were measured with a calibrated eyepiece through the microscope. |
The source is not mentioned. In the abstract, on page 5 (8ff), one can read: |
| [14.] Aeh/Fragment 013 01 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 13, Zeilen: 1-7 |
Quelle: Wernsmann et al 2006 Seite(n): 1104, Zeilen: l.col: 22-31 |
|---|---|
| A glass electrode filled with 2 m KCl was fixed in a special holder connected with plastic tube to a pressure injector and the tip inserted into the temporal neocortical slices (layer III). A high-pressure pulse was applied to inject in the tissue an amount of K+ sufficient to induce CSD (tip diameter, 2 μm; injection pressure, 0.5–1.0 bar applied for 200–300 ms, two separate injections, 1–3 nL per pulse, 2–5 mm apart from nearby recording electrodes). CSD were evaluated with respect to their amplitudes, duration and velocity rates. Duration of DC potential fluctuation width was measured at its half-maximal amplitude. | A glass electrode filled with 2 m KCl was fixed in a special holder connected with plastic tube to a pressure injector and the tip inserted into the temporal neocortical slices (layer I–II) and hippocampal tissues (CA1 area). A high-pressure pulse was applied to inject in the tissue an amount of K+ sufficient to induce SD (tip diameter, 2 μm; injection pressure, 0.5–1.0 bar applied for 200–300 ms, up to two separate injections, 1–3 nL per pulse, 2–5 mm apart from nearby recording electrodes). SD were evaluated with respect to their amplitudes, duration and velocity rates. Duration of DC potential fluctuation width was measured at its half-maximal amplitude. |
The source is not mentioned. |
| [15.] Aeh/Fragment 015 03 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 15, Zeilen: 3-6, 8-11 |
Quelle: Sheikh 2009 Seite(n): 14, Zeilen: 10-17 |
|---|---|
| Application of KCl in to neocortical slices elicited fluctuations of the DC potentials consisting of initially negative shifts, followed by positive waves. The triggered wave propagated first the nearby electrode, and a few seconds later the other electrode located parallel to this electrode. The amplitude and duration of negative DC deflections were 13.6 ± 0.8 mV and 102 ± 13.1 seconds, respectively, in the first electrode. The amplitude and duration of CSD recorded by the second electrode was 11.5 ± 0.6 mV and 93 ± 10.3 seconds, respectively. The velocity of SD propagation was determined by dividing the distance between two microelectrodes by the interval of DC potential shift appearances. The velocity of vertical propagation of DC fluctuation was 3.6 ± 0.2 mm/min. | Application of KCl into hippocampal slices elicited deflections of the DC potentials consisting of initially negative shifts, followed by positive waves (Fig. 1A). The amplitude and duration of negative DC deflections were 12.6 ± 0.7 mV and 93 ± 12.4 seconds, respectively. The triggered wave propagated against the flow of the superfusate and reached first the nearby electrode, and a few seconds later the other electrode located closer to the inlet of superfusate in the chamber. The velocity of SD propagation was determined by dividing the distance between two microelectrodes by the interval of DC potential shift appearances. The velocity of vertical propagation of DC fluctuation was 3.3 ± 0.1 mm/min. |
The source is not mentioned. The text has been adapted to accomodate the different set-up and results. Note that it seems that "The triggered wave propagated against the flow of the superfusate and reached first the nearby electrode" was shortened to "The triggered wave propagated first the nearby electrode", which is not grammatically correct. This could be seen as an indication, that indeed Aeh (2009) has taken text from Sheikh (2009) and not the other way round. |
| [16.] Aeh/Fragment 017 06 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 17, Zeilen: 6-16 |
Quelle: Sheikh 2009 Seite(n): 22, Zeilen: 7-19 |
|---|---|
| SD cellular correlate is a depolarization shift associated with complete breakdown of the membrane potential that is dependent on the asymmetric intra- and extracellular ion distribution and is maintained by the energy consuming work of membrane pumps. The ability of neurons to generate action potentials is lost during SD, which explains the spreading electrical silence accompanied SD under physiological conditions (Somjen 2001). In [sic] the other hand, epileptic activity is characterized by the paroxysmal depolarization shift, a hypersynchronous network event resulting from a giant excitatory postsynaptic potential. A paroxysmal depolarization shift is the correlate of. [sic] The EPSP is presumably the consequence of synchronous activation of recurrent excitatory pathways (Jonston and Brown, 1984). SD is accompanied by a very large increase in [K+]o. The increase in [K+]o is accompanied by a precipitous drop in [Cl-]o, [Na+]o, and [Ca2+]o. | SD cellular correlate is a depolarization shift associated with complete breakdown of the membrane potential that is dependent on the asymmetric intra-/extracellular ion distribution and is maintained by the energy consuming work of membrane pumps. The ability of neurons to generate action potentials is lost during SD, which explains the spreading electrical silence (depression of electrocorticographic activity) accompanied SD under physiological conditions (Somjen 2001). In [sic] the other hand, epileptic activity is characterized by the paroxysmal depolarization shift, a hypersynchronous network event resulting from a giant excitatory postsynaptic potential. A paroxysmal depolarization shift is the correlate of. [sic] The EPSP is presumably the consequence of synchronous activation of recurrent excitatory pathways (Jonston and Brown, 1984). [...] SD is accompanied by a very large increase in [K+]o. The increase in [K+]o is accompanied by a precipitous drop in [Cl-]o, [Na+]o, and [Ca2+]o. |
The source is not mentioned. Note that also errors have been copied from the source: "In the other hand", The truncated sentence: "A paroxysmal depolarization shift is the correlate of." |
| [17.] Aeh/Fragment 017 17 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 17, Zeilen: 17-23 |
Quelle: Gorji 2001 Seite(n): 38, Zeilen: l.col: 20ff |
|---|---|
| It was postulated that the excitatory amino acid glutamate plays a role in the chain of events triggering SD (Bures et al., 1974). The neocortex releases excitatory amino acids including glutamate and aspartate, to the extracellular space during CSD (Van Harreveld and Kooiman, 1965). Subsequently it has been shown that the triggering of SD requires activation of the NMDA subtype of glutamate receptor in rat cerebral cortex (Bures et al., 1974), in chick retina (Seeling, 1993) and in human neocortical tissues (Gorji et al., 2001). Other glutamate subreceptor (AMPA, kainate and quisqualate) can induce SD but their initiation inhibited by [NMDA receptor antagonist (Lauritzen et al., 1988).]
Bures J., Buresova O., Krivanek J. In: The Mechanisms and Applications of Leao’s Spreading Depression of Electroencephalographic Activity, Academic Press, New York (1974). Gorji A, Scheller D, Straub H, Tegtmeier F, Köhling R, Höhling JM, Tuxhorn I, Ebner A, Wolf P, Werner Panneck H, Oppel F, Speckmann EJ. Spreading depression in human neocortical slices. Brain Res. 2001;906:74-83. Lauritzen M, Rice ME, Okada Y, Nicholson C. Quisqualate, kainate and NMDA can initiate spreading depression in the turtle cerebellum. Brain Res. 1988; 475:317–327. Seelig MS, Interrelationship of magnesium and estrogen in cardiovascular and bone disorders, eclampsia, migraine and premenstrual syndrome. J. Am. Coll. Nutr. 1993;12:442–458. Van Harreveld A, Kooiman M. Amino acid release from the cerebral cortex during spreading depression and asphyxiation. J. Neurochem. 1965;12:431–439. |
It was postulated that the excitatory amino acid glutamate plays a role in the chain of events triggering SD [442,443]. The brain cortex releases excitatory amino acids including glutamate and aspartate, to the extracellular space during CSD [447]. Subsequently it has been shown that the triggering of SD requires activation of the NMDA subtype of glutamate receptor in rat cerebral cortex [147,261], in chick retina [380] and in human neocortical tissues [149]. Other glutamate receptor subtypes (AMPA, kainate and quisqualate) can induce SD but their effects inhibited by NMDA receptor antagonist [241,383].
[147] N.A. Gorelova, V.I. Koroleva, T. Amemori, V. Pavlik, J. Bures, Ketamine blockade of cortical spreading depression in rats, Electroencephalogr. Clin. Neurophysiol. 66 (1987) 440-447. [149] A. Gorji, D. Scheller, H. Straub, F. Tegtmeier, A. Ebnen, P. Wolf, H.W. Panneck, F. Oppel, E.-J. Speckmann, R. Kohling, J. Hohling, I. Tuxhorn, Spreading depression in neocortical human slices, Brain Res. 906 (2001) 74-83. [241] M. Lauritzen, M.E. Rice, Y. Okada, C. Nicholson, Quisqualate, kainate and NMDA can initiate spreading depression in the turtle cerebellum, Brain Res. 475 (1988) 317-327. [261] R. Marrannes, R. Willems, E. De Prins, A. Wauquier, Evidence for a role of the N-metPyl-D-aspartate (NMDA) receptor in cortical spreading depression in the rat, Brain Res. 457 (1988) 226-240. [380] M.S. Seelig, Interrelationship of magnesium and estrogen in cardiovascular and bone disorders, eclampsia, migraine and premenstrual syndrome, J. Am. Coll. Nutr. 12 (1993) 442-458. [383] M.J. Sheardown, The triggering of spreading depression in the chicken retina: a pharmacological study, Brain Res. 607 (1993) 189-194. [442] A. Van Harreveld, Compounds in brain extracts causing spreading depression of cerebral cortical activity and contraction of crustacean muscle, J. Neurochem. 3 (1959) 300-315. [443] A. Van Harreveld, E. Fifkova, Glutamate release from the retina during spreading depression, J. Neurobiol. 2 (1970) 13-29. [447] A. Van Harreveld, M. Kooiman, Amino acid release from the cerebral cortex during spreading depression and asphyxiation, J. Neurochem. 12 (1965) 431-439. |
The source is not mentioned. The correct Gorji et al. paper appears to have the title: "Spreading depression in human neocortical slices"; the other seems not to exist. |
| [18.] Aeh/Fragment 018 01 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 18, Zeilen: 1ff (complete) |
Quelle: Gorji 2001 Seite(n): 38, Zeilen: l.col: 30ff |
|---|---|
| This suggests that glutamate evokes SD via an action at NMDA receptors.
Migraine sufferers (notably with aura) have substantially higher plasma glutamate and aspartate levels than controls and tension headache patients between attacks. During migraine attacks, glutamate and to a lesser extent aspartate levels are even further increased which show a defective cellular reuptake mechanism for these excitatory amino acids in migraineurs (Ferrari et al., 1990). Another study showed that migraine patients during attacks had higher CSF concentrations of glutamic acid than in controls (Martinez et al., 1993). Aura symptoms cause severe disability over many hours to days in patients with familial hemiplegic migraine (FHM). Intranasal application of the NMDA antagonist ketamine reversibly reduced the severity and duration of neurological deficit in FHM (Kaube et al., 2000). Subcutaneous administration of ketamine produced a marked relief of pain both as an acute treatment and as a prophylactic therapy in migraineurs (Nicolodi and Sicuteri, 1995). In addition, it was suggested that NMDA-mediated transmission is involved in nociceptive trigeminovascular transmission within the trigeminocervical complex in cats (Goadsby and Classey, 2000). The NMDA receptor antagonist MK-801 reduces capsaicin-induced c-fos expression within rat trigeminal nucleus caudalis suggest that NMDA receptors provide a potential therapeutic target for cephalic pain (e.g., migraine) due to trigeminovascular activation from meningeal afferents (Mitsikostas et al., 1989). 5-HT1A receptor stimulation suppresses NMDA receptor-mediated synaptic excitation in the rat visual cortex (Edagawa et al., 1999). NMDA receptor blockade as well as activation of the 5-HT1A receptor attenuates the properties of KCl-induced SD in parietal cortical slices of adult rats (Kruger et al., 1999). Migraine patients have a greater cerebral 5-HT1A hypersensitivity and several anti-migraine agents exhibit marked 5-HT1A receptor activity (Leone et al., 1998). Edagawa Y, Saito H, Abe K. Stimulation of the 5-HT1A receptor selectively suppresses NMDA receptor-mediated synaptic excitation in the rat visual cortex. Brain Res. 1999;827:225–228. Ferrari MD, Odink J, Bos KD, Malessy MJ, Bruyn GW. Neuroexcitatory plasma amino acids are elevated in migraine. Neurology 1990; 40:1582–1586. Goadsby PJ, Classey JD. Glutamatergic transmission in the trigeminal nucleus assessed with local blood flow. Brain Res. 2000;875:119–124. Kaube H, Herzog J, Kaufer T, Dichgans M, Diener HC. Aura in some patients with familial hemiplegic migraine can be stopped by intranasal ketamine. Neurology 2000;55:139–141. Kruger H, Heinemann U, Luhmann HJ. Effects of ionotropic glutamate receptor blockade and 5-HT1A receptor activation on spreading depression in rat neocortical slices. Neuroreport 1999;10:2651–2656. Leone M, Attanasio A, Croci D, Ferraris A, D’Amico D, Grazzi L, Nespolo A,. Bussone G. 5- HT1A receptor hypersensitivity in migraine is suggested by the m-chlorophenylpiperazine test. Neuroreport 1998;9:2605–2608. Martinez F, Castillo J, Rodriguez JR, Leira R, Noya M. Neuroexcitatory amino acid levels in plasma and cerebrospinal fluid during migraine attacks. Cephalalgia 1993;13:89–93. Mitsikostas DD, Sanchez del Rio M, Waeber C, Moskowitz MA, Cutrer F. The NMDA receptor antagonist MK-801 reduces capsaicin-induced c-fos expression within rat trigeminal nucleus caudalis. Pain 1998;76:239–248. Nicolodi M, Sicuteri F. Exploration of NMDA receptors in migraine: therapeutic and theoretic implications. Int. J. Clin. Pharmacol. Res. 1995; 15:181–189. |
This suggests that glutamate evokes SD via an action at NMDA receptors.
Migraine sufferers (notably with aura) have substantially higher plasma glutamate and aspartate levels than controls and tension headache patients between attacks. During migraine attacks, glutamate and to a lesser extent aspartate levels are even further increased which show a defective cellular reuptake mechanism for these excitatory amino acids in migraineurs [125]. Another study showed that migraine patients during attacks had higher CSF concentrations of glutamic acid than in controls [263]. Aura symptoms cause severe disability over many hours to days in patients with familial hemiplegic migraine (FHM). Intranasal application of the NMDA antagonist ketamine reversibly reduced the severity and duration of neurological deficit in FHM [205]. Subcutaneous administration of ketamine produced a marked relief of pain both as an acute treatment and as a prophylactic therapy in migraineurs [307]. Furthermore, it was suggested that NMDA-mediated transmission is involved in nociceptive trigeminovascular transmission within the trigeminocervical complex in cats [140]. The NMDA receptor antagonist MK-801 reduces capsaicin-induced c-fos expression within rat trigeminal nucleus caudalis suggest that NMDA receptors provide a potential therapeutic target for cephalic pain (e.g., migraine) due to trigeminovascular activation from meningeal afferents [289]. As mentioned, migraineurs have a greater cerebral 5-HT1A hypersensitivity and several antimigraine agents exhibit marked 5-HT1A receptor activity [247,302]. 5-HT1A receptor stimulation suppresses NMDA receptor-mediated synaptic excitation in the rat visual cortex [116]. NMDA receptor blockade as well as activation of the 5-HT1A receptor attenuates the properties of KCl-induced SD in parietal cortical slices of adult rats [228]. [116] Y. Edagawa, H. Saito, K. Abe, Stimulation of the 5-HT1A receptor selectively suppresses NMDA receptor-mediated synaptic excitation in the rat visual cortex, Brain Res. 827 (1999) 225-228. [125] M.D. Ferrari, J. Odink, K.D. Bos, M.J. Malessy, G.W. Bruyn, Neuroexcitatory plasma amino acids are elevated in migraine, Neurology 40 (1990) 1582-1586. [140] PJ. Goadsby, J.D. Classey, Glutamatergic transmission in the trigeminal nucleus assessed with local blood flow, Brain Res. 875 (2000) 119-124. [205] H. Kaube, J. Herzog, T. Kaufer, M. Dichgans, H.C. Diener, Aura in some patients with familial hemiplegic migraine can be stopped by intranasal ketamine, Neurology 55 (2000) 139-141. [228] H. Kruger, U. Heinemann, H.J. Luhmann, Effects of ionotropic glutamate receptor blockade and 5-HT1A receptor activation on spreading depression in rat neocortical slices, Neuroreport 10 (1999) 2651-2656. [247] M. Leone, A. Attanasio, D. Croci, A. Ferraris, D. D’Amico, L. Grazzi, A. Nespolo, G. Bussone, 5-HT1A receptor hypersensitivity in migraine is suggested by the m-chlorophenylpiperazine test, Neuroreport 9 (1998) 2605-2608. [263] F. Martinez, J. Castillo, J.R. Rodriguez, R. Leira, M. Noya, Neuroexcitatory amino acid levels in plasma and cerebrospinal fluid during migraine attacks, Cephalalgia 13 (1993) 89-93. [289] D.D. Mitsikostas, M. SancPez del Rio, C. Waeber, M.A. Moskowitz, F.M. Cutrer, The NMDA receptor antagonist MK-801 reduces capsaicin-induced c-fos expression within rat trigeminal nucleus caudalis, Pain 76 (1998) 239-248. [302] A. Newman-Tancredi, C. Conte, C. Chaput, L. Verriele, V. Audinot- Bouchez, S. Lochon, G. Lavielle, M.J. Millan, Agonist activity of antimigraine drugs at recombinant human 5-HT1A receptors: potential implications for prophylactic and acute therapy, Naunyn Schmtrerbrrg’s ArcP. Pharmacol. 355 (1997) 682-688. [307] M. Nicolodi, F. Sicuteri, Exploration of NMDA receptors in migraine: therapeutic and theoretic implications, Int. J. Clin. Pharmacol. Res. 15 (1995) 181-189. |
The source is not mentioned. |
| [19.] Aeh/Fragment 019 01 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 19, Zeilen: 1-15 |
Quelle: Gorji 2001 Seite(n): 38, Zeilen: r.col: 14ff |
|---|---|
| Magnesium is known to block the NMDA receptor. The available evidence suggests that up to 50% of patients during an acute migraine attack have lowered levels of ionized magnesium (Ramadan et al., 1999). Erythrocytes and lymphocytes magnesium concentrations are significantly lower in migraine patients as compared to controls (James et al., 2000). Infusion of magnesium results in a rapid and sustained relief of an acute migraine in such patients (Mauskop et al., 1995). However, it should be mentioned that magnesium concentration has an effect on serotonin receptors, nitric oxide synthesis and release, and a variety of other migraine-related receptors and neurotransmitters (Mauskop and Altura, 1998). It is well known that reduced magnesium facilitates the development of SD in animal models and human tissues (Mody et al., 1987). Intravenous magnesium as well as the NMDA receptor antagonist MK-801 significantly reduced the frequency of SD evoked by cortical KCl application in rats. On the other hand, SD affects magnesium metabolism in the central nervous system (van der Hel et al., 1998). The magnesium sulphate anesthesia was considerably shortened approximately four times by CSD in functional decorticated mice in comparison with control animals (Bohdanecky and Necina 1963).
Bohdanecky Z, Necina J. Course of some pharmacological tests in functionally decorticated animals. Physiol. Bohemoslov. 1963;12:55–61. Mauskop A, Altura BM. Role of magnesium in the pathogenesis and treatment of migraines. Clin. Neurosci. 1998;5:24–27. Mauskop A, Altura BT, Cracco RQ, Altura BM. Intravenous magnesium sulphate relieves migraine attacks in patients with low serum ionized magnesium levels: a pilot study. Clin. Sci. 1995;89:633–636. Mody I, Lambert JD, Heinemann U. Low extracellular magnesium induces epileptiform activity and spreading depression in rat hippocampal slices. J. Neurophysiol. 1987;57:869– 888. Ramadan NM, Halvorson H, Vande-Linde A, Levine SR, Helpern JA, Welch KM. Low brain magnesium in migraine. Headache 1989;29:590–593. van der Hel WS, van den Bergh WM, Nicolay K, Tulleken KA, Dijkhuizen RM. Suppression of cortical spreading depressions after magnesium treatment in the rat. Neuroreport 1998;9:2179–2182. |
Magnesium is known to block the NMDA receptor [271]. The available evidence suggests that up to 50% of patients during an acute migraine attack have lowered levels of ionized magnesium [348,455]. Erythrocytes and lymphocytes magnesium concentrations are significantly lower in migraine patients as compared to controls [426]. Infusion of magnesium results in a rapid and sustained relief of an acute migraine in such patients [270]. However, it should be mentioned that magnesium concentration has an effect on serotonin receptors, nitric oxide synthesis and release, and a variety of other migraine-related receptors and neurotransmitters [269]. It is well known that reduced magnesium facilitates the development of SD in animal models and human tissues [16,291]. Intravenous magnesium as well as the NMDA receptor antagonist MK-801 significantly reduced the frequency of SD evoked by cortical KCl application in rats [439]. On the other hand, SD affects magnesium metabolism in the central nervous system. The magnesium sulphate anesthesia was considerably shortened approximately four times by CSD in functional decorticated mice in comparison with control animals [35].
[16] M. Avoli, C. Drapeau, J. Louvel, R. Pumain, A. Olivier, J.G. Villemure, Epileptiform activity induced by low extracellular magnesium in the human cortex maintained in vitro, Ann. Neurol. 30 (1991) 589-596. [35] Z. Bohdanecky, J. Necina, Course of some pharmacological tests in functionally decorticated animals, Physiol. Bohemoslov. 12 (1963) 55-61. [269] A. Mauskop, B.M. Altura, Role of magnesium in the pathogenesis and treatment of migraines, Clin. Neurosci. 5 (1998) 24-27. [270] A. Mauskop, B.T. Altura, R.Q. Cracco, B.M. Altura, Intravenous magnesium sulphate relieves migraine attacks in patients with low serum ionized magnesium levels: a pilot study, Clin. Sci. 89 (1995) 633-636. [271] M.L. Mayer, G.L. Westbrook, PB. Guthrie, Voltage-dependent block by Mg21 of NMDA responses in spinal cord neurones, Nature 309 (1984) 261-263. [291] I. Mody, J.D. Lambert, U. Heinemann, Low extracellular magnesium induces epileptiform activity and spreading depression in rat hippocampal slices, J. NeuropPysiol. 57 (1987) 869-888. [348] N.M. Ramadan, H. Halvorson, A. Vande-Linde, S.R. Levine, J.A. Helpern, K.M. Welch, Low brain magnesium in migraine, Headache 29 (1989) 590-593. [426] J. Thomas, J.M. Millot, S. Sebille et al., Free and total magnesium in lymphocytes of migraine patients: effect of magnesium-rich mineral water intake, Clin. Chim. Acta 295 (2000) 63-75. [439] W.S. van der Hel, W.M. van den Bergh, K. Nicolay, K.A. Tulleken, R.M. Dijkhuizen, Suppression of cortical spreading depressions after magnesium treatment in the rat, Neuroreport 9 (1998) 2179-2182. [455] K.M. Welch, G.L. Barkley, N. Tepley, N.M. Ramadan, Central neurogenic mechanisms of migraine, Neurology 43 (1993) S21- S25. |
The source is not mentioned. Note: there is no entry "James et al., 2000" in the bibliography. |
| [20.] Aeh/Fragment 019 19 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 19, Zeilen: 19-25 |
Quelle: Sheikh 2009 Seite(n): 23, 24, Zeilen: 23: 24ff - 24: 1-5 |
|---|---|
| Increasing of potassium concentration may further impairs GABA mediated inhibition and leads to appearance of ictaform burst discharges. Higashima et al. (1996) have shown that activation of GABAergic mechanisms is necessary for the generation of afterdischarges recorded in hippocampal slices after electrical stimuli. Experimental and computational data obtained by Traub et al. (1996) also suggest a role played by GABAA-mediated depolarizing conductance in the epileptiform synchronization that occurs in some models of epileptiform discharge (in [particular that induced by 4AP application).] | Increasing of potassium concentration may further impairs GABA mediated inhibition and leads to appearance of ictaform burst discharges. Higashima et al. (1996) have shown that
[Seite 24] activation of GABAergic mechanisms is necessary for the generation of afterdischarges recorded in hippocampal slices after electrical stimuli. Experimental and computational data obtained by Traub et al. (1996) also suggest a role played by GABAA-mediated depolarizing conductance in the epileptiform synchronization that occurs in some models of epileptiform discharge (in particular that induced by 4AP application). |
The source is not mentioned. |
| [21.] Aeh/Fragment 020 01 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 20, Zeilen: 1-9 |
Quelle: Sheikh 2009 Seite(n): 24, Zeilen: 5-13 |
|---|---|
| GABAergic inhibitory networks can also synchronize principal cells in the neocortex and hippocampus (Cobb et al. 1995). Higashima et al. (1996) have shown that activation of GABAergic mechanisms is necessary for the generation of afterdischarges recorded in hippocampal slices after electrical stimuli. Experimental and computational data obtained by Traub et al. (1996) also suggest a role played by GABAA-mediated depolarizing conductance in the epileptiform synchronization that occurs in some models of epileptiform discharge (in particular that induced by 4AP application). GABAergic inhibitory networks can also synchronize principal cells in the neocortex and hippocampus (Cobb et al. 1995). | GABAergic inhibitory networks can also synchronize principal cells in the neocortex and hippocampus (Cobb et al. 1995). Higashima et al. (1996) have shown that activation of GABAergic mechanisms is necessary for the generation of afterdischarges recorded in hippocampal slices after electrical stimuli. Experimental and computational data obtained by Traub et al. (1996) also suggest a role played by GABAA-mediated depolarizing conductance in the epileptiform synchronization that occurs in some models of epileptiform discharge (in particular that induced by 4AP application). GABAergic inhibitory networks can also synchronize principal cells in the neocortex and hippocampus (Cobb et al. 1995). |
The source is not mentioned. |
Quellen
| [1.] Quelle:Aeh/Herreras et al 1994 |
| Autor | Oscar Herreras, Carlota Large, José M. Ibarz, George G. Somien, Rafael Martin del Rio |
| Titel | Role of Neuronal Synchronizing Mechanisms in the Propagation of Spreading Depression in the in vivo Hippocampus |
| Zeitschrift | The Journal of Neuroscience |
| Ausgabe | 14 |
| Datum | November 1994 |
| Nummer | 11 |
| Seiten | 7087-7098 |
| URL | http://www.jneurosci.org/content/14/11/7087.long |
Literaturverz. |
no |
| Fußnoten | no |
| [2.] Quelle:Aeh/Gorji et al 2003 |
| Autor | A. Gorji, D. Scheller, E-J. Speckmann |
| Titel | The Lateral Spread of Epileptiform Discharges in Rat Neocortical Slices: Effect of Focal Phencyclidine Application |
| Zeitschrift | Pharmacopsychiatry |
| Verlag | Georg Thieme Verlag |
| Ausgabe | 36 |
| Jahr | 2003 |
| Seiten | 113-120 |
| ISSN | 0176-3679 |
| URL | https://www.thieme-connect.com/DOI/DOI?10.1055/s-2003-39986 |
Literaturverz. |
no |
| Fußnoten | no |
| [3.] Quelle:Aeh/Gorji 2001 |
| Autor | Ali Gorji |
| Titel | Spreading depression: a review of the clinical relevance |
| Zeitschrift | Brain Research Reviews |
| Verlag | Elsevier |
| Ausgabe | 38 |
| Jahr | 2001 |
| Seiten | 33-60 |
| URL | http://www.sciencedirect.com/science/article/pii/S0165017301000819 |
Literaturverz. |
yes |
| Fußnoten | yes |
| [4.] Quelle:Aeh/Smith et al 2006 |
| Autor | Justin M. Smith, Daniel P. Bradley, Michael F. James, Christopher L.-H. Huang |
| Titel | Physiological studies of cortical spreading depression |
| Zeitschrift | Biological Reviews |
| Herausgeber | Cambridge Philosophical Society |
| Ausgabe | 81 |
| Jahr | 2006 |
| Seiten | 457–481 |
| DOI | 10.1017/S1464793106007081 |
| URL | http://onlinelibrary.wiley.com/doi/10.1111/j.1469-185X.2006.tb00214.x/abstract |
Literaturverz. |
no |
| Fußnoten | no |
| [5.] Quelle:Aeh/Wernsmann et al 2006 |
| Autor | Brigitta Wernsmann, Hans-Christian Pape, Erwin-Josef Speckmann, Ali Gorji |
| Titel | Effect of cortical spreading depression on synaptic transmission of rat hippocampal tissues |
| Zeitschrift | European Journal of Neuroscience |
| Ausgabe | 23 |
| Datum | March 2006 |
| Nummer | 5 |
| Seiten | 1103–1110 |
| DOI | 10.1111/j.1460-9568.2006.04643.x |
| URL | http://onlinelibrary.wiley.com/doi/10.1111/j.1460-9568.2006.04643.x/abstract |
Literaturverz. |
yes |
| Fußnoten | yes |
| [6.] Quelle:Aeh/Sheikh 2009 |
| Autor | Khalil Sheikh |
| Titel | Spreading depression triggers ictal activity in disinhibited hippocampal slices |
| Ort | Münster |
| Jahr | 2009 |
| Anmerkung | Inaugural-Dissertation zur Erlangung des doctor medicinae der Medizinischen Fakultät der Westfälischen Wilhelms-Universität Münster, Tag der mündlichen Prüfung: 13.01.2009, Online publication: 05.02.2009 |
| URL | http://miami1.uni-muenster.de/servlets/DocumentServlet?id=4626 , http://d-nb.info/992877040/34 |
Literaturverz. |
no |
| Fußnoten | no |
| [7.] Quelle:Aeh/Somjen 2001 |
| Autor | George G. Somjen |
| Titel | Mechanisms of spreading depression and hypoxic spreading depression-like depolarization |
| Zeitschrift | Physiological Reviews |
| Herausgeber | American Physiological Society |
| Ausgabe | 81 |
| Jahr | 2001 |
| Nummer | 3 |
| Seiten | 1065-1096 |
| URL | http://physrev.physiology.org/content/physrev/81/3/1065.full.pdf |
Literaturverz. |
yes |
| Fußnoten | yes |