This report is based on the findings of an on-going plagiarism analysis (date: 2014-12-29). It is therefore not a final or conclusive report. It is recommended to visit the page http://de.vroniplag.wikia.com/wiki/Tim for the current state of the findings and further information.
A critical discussion of the doctoral thesis by Dr. Tiziana Masullo: Identification and characterization of Fluorescent Protein from marine organisms and potentially applications
Submitted to the Department of Environmental Sciences, Informatics and Statistics of the Ca'Foscari University of Venice, in fulfillment of the requirements for a research doctorate in Science and Technology. Coordinator: Prof. Bruno Pavoni. Supervisors: Prof. Marcantonio Bragadin, Dott.ssa Angela Cuttitta. Thesis published: Venice, 15.3.2012 → Download DSpace
The barcode displays a visualization of the pages that contain plagiarism, not the amount of plagiarism in the main text. Depending on the amount of plagiarized text there are three colors that are used:
- black: up to 50 % of the lines on the page are plagiarized
- dark red: between 50 % and 75 % of the lines on the page are plagiarized
- light red: over 75 % of the lines on the page are plagiarized.
White pages have either not yet been investigated or nothing was found. Blue pages contain matter such as the title page, the table of contents, the reference section, empty pages and appendices. These are all not included in the calculations.
The barcode only shows the current state of the investigation. This is not the final result, as each case may continue to be worked on and added to by anyone as new sources turn up. Thus, a final state does not exist.
There are 25 pages containing plagiarism.
Pages with less than 50% plagiarism
8 Pages: 072 041 023 017 024 015 069 070
Pages with between 50%-75% plagiarism
Pages with more than 75% plagiarism
14 Pages: 073 019 006 042 008 009 010 007 012 013 011 025 016 020
Prominent fragments
- Fragment 072 18, Fragment 073 01: The last paragraph of the thesis, about a page in length, is taken verbatim from the publication Shang & Dong (2008) that is nowhere mentioned in the thesis. Also twelve references to the literature are copied.
- Fragment 071 05: Here the source is given, but only for a part of the text taken from it. Remarkable is also that the source seems to be significantly misrepresented.
- Fragment 023 19: The source is given, but also two sentences starting with "Accordingly, we observed" were taken from it literally, such that the reader must (wrongly) assume that the author made observations consistent with the findings of the source.
- Fragment 024 09: An image has been taken from the Wikipedia without attribution.
Prominent sources
- Wikipedia (Snakelocks anemone): A section of the thesis has been taken verbatim from the Wikipedia that is not mentioned in the thesis anywhere. It is remarkable that in the PDF file of the thesis that is available for download, one can find invisible, but active Wikipedia links at the same places where they also appear (and are visible) in the original Wikipedia article.
Statistics
- Currently there are 34 reviewed fragments documented that are considered to be violations of citation rules. For 14 of them there is no reference given to the source used („Verschleierungen“ and „Komplettplagiate“). For 20 fragments the source is given, but the extent of the used text is not made clear („Bauernopfer“).
- The publication has 75 pages that have been analyzed. On a total of 25 of these pages violations of citation rules have been documented. This represents a percentage of 33.3%. The 75 analyzed pages break down with respect to the amount of text parallels encountered as follows:
- From these statistics an extrapolation of the amount of text of the publication under investigation that has been documented as problematic can be estimated (conservatively) as about 18% of the main part of the publication.
- In all, text was taken from 19 sources.
Illustration
The following chart illustrates the amount and the distribution of the text parallel findings. The colours show the type of plagiarism diagnosed:
- grau="Komplettplagiat" (copy & paste): the source of the text parallel is not given, the copy is verbatim.
- rot="Verschleierung" (disguised plagiarism): the source of the text parallel is not given, the copied text will be somewhat modified.
- gelb="Bauernopfer" (pawn sacrifice): the source of the text parallel is mentioned, but the extent and/or the closeness of the copy to the source is not made clear by the reference.

Definitions of plagiarism categories
The plagiarism categories used here are based on the discussion found at Wohnsdorf / Weber-Wulff: Strategien der Plagiatsbekämpfung, 2006. A complete description of the categories (in German) can be found at the VroniPlag-Wiki. In particular, the categories are:
Translation plagiarism
A translation plagiarism is based on a text in a differentlanguage than the text of the thesis and represents a word for word or literal translation. The difference between paraphrase (which is properly referenced) and translation plagiarism is sometimes seen as hard to judge.
Komplettplagiat (copy and paste)
The source of the text parallel is not given, the copy is verbatim.
Verschleierung (disguised plagiarism)
The source of the text parallel is not given, the copied text will be somewhat modified or disguised.
Bauernopfer (pawn sacrifice)
The source of the text parallel is mentioned, but the extent and/or closeness of the copying is not made clear by the reference.
Sources tabulated by plagiarism category
The following table lists all of the reviewed fragments by source (rows) and by plagiarism category (columns).
- ÜP = Translation plagiarism,
- KP = Copy and paste,
- VS = Disguised plagiarism,
- BO = Pawn sacrifice,
- KW = No opinion,
- KeinP = Not plagiarism.
| Source |
Year | ÜP |
KP |
VS |
BO |
KW |
KeinP |
∑ |
To be reviewed |
In progress |
|---|---|---|---|---|---|---|---|---|---|---|
| Andresen et al | 2005 | 0 | 2 | 0 | 1 | 1 | 0 | 4 | 0 | 0 |
| Chapleau | 2008 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 |
| Copper | 2010 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Fu | 2008 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 |
| Gurskaya | 2003 | 0 | 0 | 0 | 2 | 0 | 0 | 2 | 0 | 0 |
| Isarankura | 2009 | 0 | 0 | 2 | 1 | 1 | 0 | 4 | 0 | 0 |
| Lukyanov | 2000 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 |
| MSIP | 2010 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Nienhaus | 2006 | 0 | 0 | 0 | 5 | 0 | 0 | 5 | 0 | 0 |
| Rahimi et al | 2008 | 0 | 0 | 0 | 1 | 1 | 0 | 2 | 0 | 0 |
| Schaefer | 2007 | 0 | 0 | 0 | 2 | 0 | 0 | 2 | 0 | 0 |
| Schaefer | 2007b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Shang Dong | 2008 | 0 | 2 | 0 | 0 | 0 | 0 | 2 | 0 | 0 |
| Sumner et al | 2006 | 0 | 1 | 0 | 1 | 0 | 0 | 2 | 0 | 0 |
| Tasdemir et al | 2008 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 |
| Wang | 2007 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Wiedenmann | 2000 | 0 | 0 | 0 | 4 | 0 | 0 | 4 | 1 | 0 |
| Wikipedia Snakelocks anemone | 2012 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 0 | 0 |
| Wikipedia Conica (suborder) | 2009 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| ∑ | - | 0 | 7 | 7 | 20 | 3 | 0 | 37 | 3 | 0 |
Fragments
34 reviewed fragments
| Fragment | SeiteArbeit | ZeileArbeit | Quelle | SeiteQuelle | ZeileQuelle | Typus |
|---|---|---|---|---|---|---|
| Tim/Fragment 001 02 | 1 | 2-15 | Gurskaya 2003 | 403 | l.col.: 1 ff. | BauernOpfer |
| Tim/Fragment 001 19 | 1 | 19-26 | Wang 2007 | 60 | 4 ff. | KomplettPlagiat |
| Tim/Fragment 006 01 | 6 | 1-3, 4-15 | Nienhaus 2006 | 4211 , 4212 | 4211: l.col., 3 ff.; 4212 l.col. 11 ff. | BauernOpfer |
| Tim/Fragment 007 01 | 7 | 1 ff. (komplett) | Nienhaus 2006 | 4212, 4213 | 4212: l.col.: 30 ff.; 4213: l.col: 1ff | BauernOpfer |
| Tim/Fragment 008 01 | 8 | 1-7 | Nienhaus 2006 | 4213 | l.col: 1 ff. | BauernOpfer |
| Tim/Fragment 008 10 | 8 | 10-19 | Tasdemir et al 2008 | 613, 621 | 613: r. col: 9 ff.; 621: r.col: 17 ff. | BauernOpfer |
| Tim/Fragment 008 21 | 8 | 21-28 | Wiedenmann 2000 | 14092 | l.col, last lines | BauernOpfer |
| Tim/Fragment 009 01 | 9 | 1 ff. (entire page) | Wiedenmann 2000 | 14092 | r.col.: 8 ff. | BauernOpfer |
| Tim/Fragment 010 01 | 10 | 1 ff. (entire page) | Wiedenmann 2000 | 14092, 14093 | 14092: r.col: last lines; 14093: l.col: 1ff | BauernOpfer |
| Tim/Fragment 011 01 | 11 | 1-28 | Wiedenmann 2000 | 14095 | l.col: 1 ff. | BauernOpfer |
| Tim/Fragment 012 01 | 12 | 1-2, 3-15 | Nienhaus 2006 | 4213, 4215, 4216 | 4213: r.col: 17 ff.; 4215: r.col.: 8 ff.; 4216: l.col. 1 ff. | BauernOpfer |
| Tim/Fragment 013 01 | 13 | 1-10 | Nienhaus 2006 | 4216 | l.col.: 2 ff. | BauernOpfer |
| Tim/Fragment 013 12 | 13 | 12-15 | Andresen et al 2005 | 13070 | l.col: 27 ff. | KomplettPlagiat |
| Tim/Fragment 014 05 | 14 | 5-9 | Lukyanov 2000 | 25880 | l.col.: 21 ff. | BauernOpfer |
| Tim/Fragment 014 10 | 14 | 10-15 | Andresen et al 2005 | 13070 | 44 ff. | KomplettPlagiat |
| Tim/Fragment 014 19 | 14 | 19-24, 27-33 | Andresen et al 2005 | 13071 | r.col. 14 ff. | BauernOpfer |
| Tim/Fragment 015 11 | 15 | 11-15 | Schaefer 2007 | 540 | l.col.: 3 ff. | BauernOpfer |
| Tim/Fragment 016 01 | 16 | 1-7 | Schaefer 2007 | 540 | l.col.: 11 ff. | BauernOpfer |
| Tim/Fragment 017 15 | 17 | 15-16 | Isarankura 2009 | 353 | 17 ff. | Verschleierung |
| Tim/Fragment 019 01 | 19 | 1-2 | Chapleau 2008 | 619 | l.col.: 14 ff. | Verschleierung |
| Tim/Fragment 019 03 | 19 | 3-16 | Isarankura 2009 | 352 | abstract | BauernOpfer |
| Tim/Fragment 020 01 | 20 | 1-15 | Sumner et al 2006 | 1303, 1306 | 1303: l.col: 25 ff.; 1305: l.col: 21 ff.; 1306: r.col: 7 ff. | BauernOpfer |
| Tim/Fragment 023 03 | 23 | 3-5 | Isarankura 2009 | 353 | last lines | Verschleierung |
| Tim/Fragment 023 19 | 23 | 19-26 | Fu 2008 | abstract, p.6 | p.6: last lines | BauernOpfer |
| Tim/Fragment 024 06 | 24 | 6-11 | MSIP 2010 | 1 (online source) | - | Verschleierung |
| Tim/Fragment 024 09 | 24 | image | Wikipedia Conica (suborder) 2009 | 1 (online source) | - | KomplettPlagiat |
| Tim/Fragment 025 01 | 25 | 1 ff. (entire page) | Gurskaya 2003 | 404 | l.col. last lines | BauernOpfer |
| Tim/Fragment 041 08 | 41 | 8-10 | Wikipedia Snakelocks anemone 2012 | online source | - | Verschleierung |
| Tim/Fragment 042 02 | 42 | 2-18 | Wikipedia Snakelocks anemone 2012 | online source | - | Verschleierung |
| Tim/Fragment 069 25 | 69 | 25-32 | Copper 2010 | 1 (online source) | - | Verschleierung |
| Tim/Fragment 070 29 | 70 | 29-32 | Sumner et al 2006 | 1306 | r.col: 7 ff. | KomplettPlagiat |
| Tim/Fragment 071 05 | 71 | 5-14 | Rahimi et al 2008 | 5 | 15 ff. | BauernOpfer |
| Tim/Fragment 072 18 | 72 | 18-32 | Shang Dong 2008 | 1569 | l.col, 1 ff. | KomplettPlagiat |
| Tim/Fragment 073 01 | 73 | 1 ff. (complete page) | Shang Dong 2008 | 1569 | l.col. 21 ff. | KomplettPlagiat |
Fragments
Remark on the colouring
The colouring is automatically generated and shows text parallels. Its purpose is to facilitate the orientation of the reader, it does not, however, automatically diagnose plagiarism of any kind. In order to form a judgement about a certain text parallel one should consult the text itself.
Remark on the line numbering
When identifying a fragment with line numbers everything that contains text (except for the page header and/or footer) is counted, including headings. However, charts and tables, including their captions, are usually not counted.
34 reviewed fragments
| [1.] Tim/Fragment 001 02 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 1, Zeilen: 2-15 |
Quelle: Gurskaya 2003 Seite(n): 403, Zeilen: l.col.: 1 ff. |
|---|---|
| Green Fluorescent Protein (GFP) from the hydromedusa Aequorea victoria is intensively used in biomedical sciences (Tsien, 1998).
In nature, GFP is a component of the A. victoria bioluminescent system. Like many other marine organisms, this jellyfish can produce bright flashes in response to external stimulation. GFP is a secondary emitter that transforms blue light (460 nm), emitted by the Ca2+- dependent photoprotein aequorin, into green light (508 nm) (Johnson et al., 1962). Although GFPs similar to Aequorea GFP are found in other bioluminescent coelenterates (Chalfie, 1995), the biological significance of blue light transformation is not clear, particularly since the majority of bioluminescent animals do not possess this mechanism. The discovery of GFP-like proteins in the non-bioluminescent Anthozoa species indicates these proteins are not necessarily linked to bioluminescence (Matz et al., 1999). In fact, these organisms can produce bright flashes through GFP, as an electromagnetic radiation source with specific wavelength in response to external stimulation (Gurskaya et al., 2003). Tsien, R., 1998. Annu. Rev. Biochem. 67: 509-544. Johnson, F. H., Shimomura, O., Saiga, Y., Gershman, L. C., Reynolds, G. T., and Waters, J. R., 1962. J. Cell. Comp. Physiol. 60, 85-104 Chalfie, M., 1995. Green fluorescent protein. Photochem. Photobiol. 62, 651–656 Matz M.V., Fradkov A.F., Labas Y.A., Savitsky A.P., Zaraisky A.G., Markelov M.L., Lukyanov S.A., 1999. Nat. Biotechnol., 17:969-973. |
Green fluorescent protein (GFP) from the hydromedusa Aequorea victoria is intensively used in biomedical sciences [1]. In nature, GFP is a component of the A. victoria bioluminescent system. Like many other marine organisms, this jellyfish can produce bright flashes in response to external stimulation. GFP is a secondary emitter that transforms blue light (460 nm), emitted by the Ca2+- dependent photoprotein aequorin, into green light (508 nm) [2]. Although GFPs similar to Aequorea GFP are found in other bioluminescent coelenterates (for review see [3]), the biological significance of blue light transformation is not clear, particularly since the majority of bioluminescent animals do not possess this mechanism. The discovery of GFP-like proteins in the non-bioluminescent Anthozoa species indicates these proteins are not necessarily linked to bioluminescence [4].
1 Tsien, R. Y. (1998) The green fluorescent protein. Annu. Rev. Biochem. 67, 509–544 2 Johnson, F. H., Shimomura, O., Saiga, Y., Gershman, L. C., Reynolds, G. T. and Waters, J. R. (1962) Quantum efficiency of Cypridina luminescence, with a note on that of Aequorea. J. Cell. Comp. Physiol. 60, 85–104 3 Chalfie, M. (1995) Green fluorescent protein. Photochem. Photobiol. 62, 651–656 4 Matz,M.V.,Fradkov, A. F., Labas, Y. A., Savitsky, A. P., Zaraisky, A. G., Markelov, M. L. and Lukyanov, S. A. (1999) Fluorescent proteins from nonbioluminescent Anthozoa species. Nat. Biotechnol. 17, 969–973 |
The source is given at the end of the paragraph. It is remarkable that the only sentence the reader sensibly would expect to come from the source cannot be found in it, while the rest of the documented passage is taken from it verbatim . |
| [2.] Tim/Fragment 001 19 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 1, Zeilen: 19-26 |
Quelle: Wang 2007 Seite(n): 60, Zeilen: 4 ff. |
|---|---|
| In fact, Fluorescent Proteins (FPs) are widely used as noninvasive probes to study different biological models from individual cells to whole organisms. The use of FPs enable the tracking of every step of the protein of interest: expression, localization, movement, interaction and activity in the cell, tissue or organism. The main applications of FPs are: visualization of target-gene promoter up- and down-regulation, protein labeling, detection of protein-protein interactions, tracking protein movement and monitoring cellular parameters using FP-based fluorescent sensors. | FPs are widely used as noninvasive probes to study different biological models from individual cells to whole organisms. The use of FPs enables the tracking of every step of the protein of interest: expression, localization, movement, interaction and activity in the cell, tissue or organism. The main applications of FPs are: visualization of target-gene promoter up- and down-regulation, protein labeling, detection of protein-protein interactions, tracking protein movement and monitoring cellular parameters using FP-based fluorescent sensors. |
The source is not mentioned anywhere. |
| [3.] Tim/Fragment 006 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 6, Zeilen: 1-3, 4-15 |
Quelle: Nienhaus 2006 Seite(n): 4211 , 4212, Zeilen: 4211: l.col., 3 ff.; 4212 l.col. 11 ff. |
|---|---|
| [Nienhaus et al. (2006) described the quaternary structure and found that the asymmetric unit of AsGFP contains two identical tetramers related by a no crystallographic symmetry. They are arranged as dimers of dimers, so that two types of subunit interfaces can be distinguished; they are denoted as antiparallel (between subunits A/B] and C/D) and perpendicular (between subunits A/C and B/D) interfaces according to the mutual orientations of the main axes of the β-barrels. Both interfaces involve hydrophobic and hydrophilic interactions (fig. 1).
[...] Relatively at tertiary structure, the monomeric subunits within the tetramers are essentially identical. The overall backbone topology shows the typical 11-stranded b-barrel [sic] fold, with the central a-helix [sic] interrupted by the chromophore. The N-terminal end (residues 1–7) forms a lid on the same barrel, thereby assisting in shielding the interior of the can from the environment, whereas the C-terminal tail (residues 220–228) wraps around the other barrel in the A/B dimer. The AsGFP structure is similar to that of AvGFP. Backbone structural differences between AsGFP and AvGFP are most pronounced in the region corresponding to amino acids 138–141 (143–146 in AvGFP) and in the loop region formed by amino acids 195–206 (204–216 in AvGFP). The chromophore of AsGFP is a planar resonance system formed autocatalytically by residues Gln63, Tyr64, and Gly65 (fig. 2, A and B). It consists of an imidazolinone ring generated by cyclization between the Gln63-C' and the Gly65-Nα atoms and the Tyr64 [7 hydroxyphenyl group, which is made coplanar with the imidazolinone due to dehydrogenation insaturation of its Cα-Cβ bond.] |
The asymmetric unit of asFP499 contains two identical tetramers related by a noncrystallographic symmetry. They are arranged as dimers of dimers, so that two types of subunit interfaces can be distinguished; they are denoted as antiparallel (between subunits A/B and C/D) and perpendicular (between subunits A/C and B/D) interfaces according to the mutual orientations of the main axes of the β-barrels. Both interfaces involve hydrophobic and hydrophilic interactions.
[page 4212] Tertiary structure The monomeric subunits within the tetramers are essentially identical, as judged from the average root mean-square deviation (rmsd) of the Ca atoms of 0.21 A° . The overall backbone topology shows the typical 11-stranded β-barrel fold, with the central α-helix interrupted by the chromophore. The N-terminal end (residues 1–7) forms a lid on the same barrel, thereby assisting in shielding the interior of the can from the environment, whereas the C-terminal tail (residues 220–228) wraps around the other barrel in the A/B dimer. The asFP499 structure is similar to that of avGFP, with an rmsd of the Ca atoms of 1.15A° . Backbone structural differences between asFP499 and avGFP are most pronounced in the region corresponding to amino acids 138–141 (143–146 in avGFP) and in the loop region formed by amino acids 195–206 (204–216 in avGFP). The chromophore and its environment The chromophore of asFP499 is a planar resonance system formed autocatalytically by residues Gln63, Tyr64, and Gly65 (Fig. 1, A and B). It consists of an imidazolinone ring generated by cyclization between the Gln63-C9 and the Gly65-Nα atoms and the Tyr64 hydroxyphenyl group, which is made coplanar with the imidazolinone due to dehydrogenation insaturation of itsCα-Cβ bond. |
The source is mentioned once in the beginning. However, it does not become clear that the entire page (except the figure 1) is copied and that the copy is almost literal. |
| [4.] Tim/Fragment 007 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 7, Zeilen: 1 ff. (komplett) |
Quelle: Nienhaus 2006 Seite(n): 4212, 4213, Zeilen: 4212: l.col.: 30 ff.; 4213: l.col: 1ff |
|---|---|
| [It consists of an imidazolinone ring generated by cyclization between the Gln63-C' and the Gly65-Nα atoms and the Tyr64] hydroxyphenyl group, which is made coplanar with the imidazolinone due to dehydrogenation insaturation of its Cα-Cβ bond. Whereas there is a glutamine in the first position of the tripeptide instead of the serine in AvGFP, the AsGFP chromophore is essentially identical to that of AvGFP, including the cis configuration at the Tyr64-Cβ, which is encountered more frequently than the trans conformation.
Fig. 2 (A) Top and (B) side view of the electron density map of the AsGFP chromophore and its environment, contoured at 1.2 s. (C) Close-up of the AsGFP chromophore (green, carbon; red, oxygen; blue, nitrogen) and surrounding residues (black, carbon; red, oxygen; blue, nitrogen). The backbone structures are plotted as lines; the side chains are accentuated. Water molecules are plotted as red spheres. Hydrogen bonds are represented by dashed lines. Distances are given in angstroms. (Nienhaus et al., 2006) The chromophore of AsGFP is tightly encased within the β-barrel by a hydrogen bond network involving polar and charged residues and altogether 10 structural waters within a distance of 5 Angstrom from the imidazolinone oxygen. |
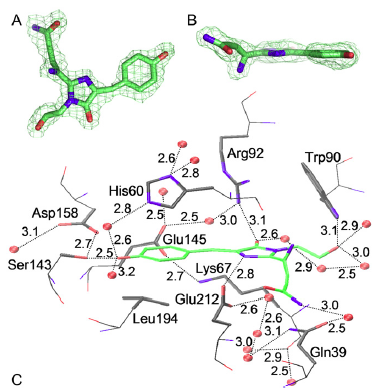
Figure 1 (A) Top and (B) side view of the electron density map of the asFP499 chromophore and its environment, contoured at 1.2 s. (C) Close-up of the asFP499 chromophore (green, carbon; red, oxygen; blue, nitrogen) and surrounding residues (black, carbon; red, oxygen; blue, nitrogen). The backbone structures are plotted as lines; the side chains are accentuated. Water molecules are plotted as red spheres. Hydrogen bonds are represented by dashed lines. Distances are given in angstroms. It consists of an imidazolinone ring generated by cyclization between the Gln63-C´ and the Gly65-Nα atoms and the Tyr64 hydroxyphenyl group, which is made coplanar with the imidazolinone due to dehydrogenation insaturation of its Cα-Cβ bond. Whereas there is a glutamine in the first position of the tripeptide instead of the serine in avGFP, the asFP499 chromophore is essentially identical to that of avGFP, including the cis configuration at the Tyr64-Cβ, which is encountered more frequently than the trans conformation. The chromophore of asFP499 is tightly encased within the β-barrel by a hydrogen-bond network involving polar and charged residues and altogether 10 structural waters within a [page 4213] distance of 5 A° from the imidazolinone oxygen. |

The source is given for the figure and its caption, but not for the remainder of the text. |
| [5.] Tim/Fragment 008 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 8, Zeilen: 1-7 |
Quelle: Nienhaus 2006 Seite(n): 4213, Zeilen: l.col: 1 ff. |
|---|---|
| Figure 2 C displays the chromophore cage, with potential hydrogen-bond interactions represented by dashed lines. The highly conserved residues Arg92 and Glu212 have been implicated as being crucially involved in the mechanism of autocatalytic chromophore formation (Ormo et al., 1996; Sniegowski et al., 2005; Wood et al., 2005). The Arg92 guanidinium group hydrogen bonds to the Tyr64-derived carbonyl oxygen, whereas Glu212 is positioned within hydrogen-bonding distance to the heterocyclic ring nitrogen. | Fig. 1 C displays the chromophore cage, with potential hydrogen-bond interactions represented by dashed lines. [...] The highly conserved residues Arg92 and Glu212 have been implicated as being crucially involved in the mechanism of autocatalytic chromophore formation (6,40,41). The Arg92 guanidinium group hydrogen bonds to the Tyr64-derived carbonyl oxygen, whereas Glu212 is positioned within hydrogen-bonding distance to the heterocyclic ring nitrogen, as has been observed earlier for yellow avGFP variants (42)
6. Ormo¨ , M., A. B. Cubitt, K. Kallio, L. A. Gross, R. Y. Tsien, and S. J. Remington. 1996. Crystal structure of the Aequorea victoria green fluorescent protein. Science. 273:1392–1395. 40. Sniegowski, J. A., J. W. Lappe, H. N. Patel, H. A. Huffman, and R. M. Wachter. 2005. Base catalysis of chromophore formation in Arg96 and Glu222 variants of green fluorescent protein. J. Biol. Chem. 280: 26248–26255. 41. Wood, T. I., D. P. Barondeau, C. Hitomi, C. J. Kassmann, J. A. Tainer, and E. D. Getzoff. 2005. Defining the role of arginine 96 in green fluorescent protein fluorophore biosynthesis. Biochemistry. 44:16211– 16220. 42. Wachter, R. M., M. A. Elsliger, K. Kallio, G. T. Hanson, and S. J. Remington. 1998. Structural basis of spectral shifts in the yellow-emission variants of green fluorescent protein. Structure. 6: 1267–1277. |
The source is given the previous page to reference the figure that is described here. Nothing indicates, however, that this section is also taken from the source. |
| [6.] Tim/Fragment 008 10 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 8, Zeilen: 10-19 |
Quelle: Tasdemir et al 2008 Seite(n): 613, 621, Zeilen: 613: r. col: 9 ff.; 621: r.col: 17 ff. |
|---|---|
| Contrary to monomeric AvGFP, all known Anthozoa GFP-like proteins reported to date form oligomers and exist mostly as tetramers (Baird et al., 2000; Mizuno et al., 2001; Wiedenmann et al., 2002; Shagin et al., 2004). The oligomerisation does not impair their application as reporter genes, selection markers or biosensors, but limits their use as fusion tags to study, for instance, protein localization and dynamics in living cells (Baird et al., 2000; Lauf et al., 2001; Mizuno et al., 2001). A possible solution to this problem is the genetic engineering of the fluorescent protein of interest by mutagenesis, creating mutants that form functional monomeric variants. For example, Tasdemir et al. (2008) converted the tetrameric AsGFP into dimeric and monomeric variants guided by a rational strategy of sequence alignments. | Contrary to monomeric AvGFP, all known Anthozoa GFP-like proteins reported to date form oligomers and exist mostly as tetramers (Baird et al., 2000; Vrzheshch et al., 2000; Mizuno et al., 2001; Wiedenmann et al., 2002; Shagin et al., 2004). The oligomerisation does not impair their application as reporter genes, selection markers or biosensors, but limits their use as fusion tags to study, for instance, protein localisation and dynamics in living cells (Baird et al., 2000; Lauf et al., 2001; Mizuno et al., 2001). A possible solution to this problem is the genetic engineering of the fluorescent protein (FP) of interest by mutagenesis, creating mutants that form functional monomeric variants.
[page 621] In summary, the present investigations led to a successful conversion of tetrameric AsGFP499 into dimeric and monomeric variants guided by a rational strategy of sequence alignments. |
The source is mentioned, but as an illustration ("For example [...]") of the previously said and not as a reference of a source for it. |
| [7.] Tim/Fragment 008 21 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 8, Zeilen: 21-28 |
Quelle: Wiedenmann 2000 Seite(n): 14092, Zeilen: l.col, last lines |
|---|---|
| Localization.
Because of their content of protein pigments, the tentacles of A. sulcata var. rufescens exhibit three hues under daylight conditions. The upper side is green, the underside is orange, and the tips of tentacles show a vivid reddish color (fig. 3A). The green and the orange pigments exhibit a bright fluorescence under irradiation with UV light at 366 nm (fig. 3B). The reddish protein of the tips is nonfluorescent. The protein pigments all are located in the ectoderm of the tentacles (fig. 3C). |
Localization. Because of their content of protein pigments, the tentacles of A. sulcata var. rufescens exhibit three hues under daylight conditions. The upper side is green, the underside is orange, and the tips of tentacles show a vivid reddish color (Fig. 1A). The green and the orange pigments exhibit a bright fluorescence under irradiation with UV light at 366 nm (Fig. 1B). In a few specimens, two opposite spots at the mouth and the verrucae of the column also fluoresce in orange. The reddish protein of the tips is nonfluorescent. The protein pigments all are located in the ectoderm of the tentacles (Fig. 1C). |
The source is mentioned on the following pages, but there is no indication that this passage is also taken from it. |
| [8.] Tim/Fragment 009 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 9, Zeilen: 1 ff. (entire page) |
Quelle: Wiedenmann 2000 Seite(n): 14092, Zeilen: r.col.: 8 ff. |
|---|---|
Fig. 3. Pigments of A. sulcata var. rufescens. (A) Localization of GFPs in the upper side, orange fluorescent protein in the underside, and reddish protein in the tips of tentacles under daylight (scale bar: 1 cm) and UV (366 nm) (B). (C) Kryo-section (20 mm) of a tentacle fixed in seawatery4% paraformaldehyde irradiated with UV (365 nm) shows the ectodermal localization of the fluorescent proteins. The orange fluorescent protein located in the underside of the tentacle shows a yellow-shifted emission. This shift most likely is induced by the fixation. The red fluorescence of the entoderm is produced by chlorophyll of zooxanthellae (scale bar: 0.5 mm). (Wiedenmann et al., 2000). Spectral properties. The spectral properties are summarized in table 1 (appendixes) (Wiedenmann et al., 2000). The green fluorescence of partially purified protein solutions is characterized by an excitation spectrum with two maxima at 480 nm and 511 nm and a shoulder around 400 nm. A region of high excitation is found in the UV spectrum. The emission spectrum shows two maxima at 499 nm and 522 nm. The ratio between the emission maxima varies in dependence of the excitation wavelength. This points to the existence of two different forms of GFP in the tissue with excitation/emission maxima at 480 nm/499 nm and 511 nm/522 nm. The orange fluorescence distinguishes itself by three excitation maxima at 278 nm, 337 nm, and 574 nm and a single emission peak at 595 nm. In the visible region the maximum of absorption of the nonfluorescent red protein is at 562 nm. According to the nomenclature introduced by Matz et al. (1999) the fluorescent proteins from A. sulcata var. rufescens were named asFP499, asFP522, and asFP595. The nonfluorescent red protein from A. sulcata var. rufescens was named asCP562. In this context, CP stands for colored protein and the number identifies the major absorption maximum. |
Spectral properties. The spectral properties are summarized in Table 1. The green fluorescence of partially purified protein solutions is characterized by an excitation spectrum with two maxima at 480 nm and 511 nm and a shoulder around 400 nm. A region of high excitation is found in the UV spectrum. The emission spectrum shows two maxima at 499 nm and 522 nm (Fig. 2A). The ratio between the emission maxima varies in dependence of the excitation wavelength. Such a variance also is found in samples derived from different specimens (data not shown). This points to the existence of two different forms of GFP in the tissue with excitation/emission maxima at 480 nm/499 nm and 511 nm/522 nm. The orange fluorescence distinguishes itself by three excitation maxima at 278 nm, 337 nm, and 574 nm and a single emission peak at 595 nm (Fig. 2A). In the visible region the maximum of absorption of the nonfluorescent red protein is at 562 nm (Fig. 2C).
According to the nomenclature introduced by Matz et al. (22) the fluorescent proteins from A. sulcata var. rufescens were named asFP499, asFP522, and asFP595. The nonfluorescent red protein from A. sulcata var. rufescens was named asCP562. In this context, CP stands for colored protein and the number identifies the major absorption maximum.
Fig. 1. Pigments of A. sulcata var. rufescens. (A) Localization of GFPs in the upper side, orange fluorescent protein in the underside, and reddish protein in the tips of tentacles under daylight (scale bar: 1 cm) and UV (366 nm) (B). (C) Kryo-section (20 mm) of a tentacle fixed in seawatery4% paraformaldehyde irradiated with UV (365 nm) shows the ectodermal localization of the fluorescent proteins. The orange fluorescent protein located in the underside of the tentacle shows a yellow-shifted emission. This shift most likely is induced by the fixation. The red fluorescence of the entoderm is produced by chlorophyll of zooxanthellae (scale bar: 0.5 mm). |
Source is mentioned twice: once to reference the image and its caption and once for table 1. It does not become clear to the reader, however, that the entire page is taken from the source. |
| [9.] Tim/Fragment 010 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 10, Zeilen: 1 ff. (entire page) |
Quelle: Wiedenmann 2000 Seite(n): 14092, 14093, Zeilen: 14092: r.col: last lines; 14093: l.col: 1ff |
|---|---|
| Molecular masses.
Wiedenmann et al. (2000) purified all proteins to near homogeneity. Both AsGFP 499/522 could not be separated by the purification steps. The molecular masses, determined by SDS-PAGE, are 26.2 kDA for the mixed fraction of AsGFP499/522 and 19.1 kDa for both asFP595 and asCP562. In gel filtration experiments under physiological conditions all protein pigments show apparent molecular masses of 66 kDa. For the GFPs this indicates a natural occurrence as dimers or trimers. Under denaturing conditions these oligomers can be partially split into monomers with apparent molecular masses of 23 kDa. Molecular masses for the AsGFP499 and asCP562 cloned were 25.4 kDa and 16.5 kDa. Stability. All colored proteins show a remarkable stability after treatment with heat, detergent, chaotrop, reducing agent, and pH extremes (Table 2. appendixes). Thermostability of asFP595 fluorescence is clearly increased compared with that of AsGFP499/522. In contrast, fluorescence of AsGFP499 is more stable when the solutions are exposed to 1% SDS, 8 M urea, and pH 11. Fluorescence of AsGFP499/522 and asFP595 is also stable when the proteins are treated with 4% paraformaldehyde (Wiedenmann et al., 2000). Reversible denaturation. Wiedenmann et al. (2000) heat-denatured the partially purified extracts of the proteins asFP595 and asCP562 in the presence of 2% SDS and 10% β-mercaptoethanol. After, proteins were separated by SDS/PAGE and transferred to a nitrocellulose membrane by semidry blotting. On the membrane, the band corresponding to asFP595 appears red under daylight conditions and exhibits orange fluorescence with an emission maximum at 595 nm under UV light. The fluorescence is stable and can even be detected after passage of the renatured protein through the membrane because of extended blotting. Renaturation with full restoration of fluorescence in the presence of β-mercaptoethanol has been described for heat-denatured GFP (Surpin & Ward, 1989). Surprisingly, asCP562, which is nonfluorescent in vivo, behaves in the same manner and shows identical fluorescence as renatured asFP595. |
Molecular masses [...] All proteins were purified to near homogeneity. Both asFP499 and asFP522 could not be separated by the purification steps. The molecular masses, determined by
[page 14093] SDS/PAGE are 26.2 kDA for the mixed fraction of asFP499/ asFP522 and 19.1 kDa for both asFP595 and asCP562 (Fig. 3 A and B). In gel filtration experiments under physiological conditions all protein pigments show apparent molecular masses of 66 kDa (Fig. 3.1 and 3.3). For the GFPs this indicates a natural occurrence as dimers or trimers (Fig. 3.3). Under denaturing conditions these oligomers can be partially split into monomers with apparent molecular masses of 23 kDa (Fig. 3.4). Stable dimers also are reported for GFP from Renilla and some other pennatularians (12, 27). Stability. All colored proteins show a remarkable stability after treatment with heat, detergent, chaotrop, reducing agent, and pH extremes (Table 2). Thermostability of asFP595 fluorescence is clearly increased compared with that of asFP499/asFP522. In contrast, fluorescence of asFP499 is more stable when the solutions are exposed to 1% SDS, 8 M urea, and pH 11. Fluorescence of asFP499/asFP522 and asFP595 is also stable when the proteins are treated with 4% paraformaldehyde. Overall, the stability of the pigments from A. sulcata is comparable to that of the GFPs from Aequorea and Renilla (27–30). Reversible denaturation. Partially purified extracts of the proteins asFP595 and asCP562 were heat-denatured in the presence of 2% SDS and 10% β-mercaptoethanol. Proteins were separated by SDS/PAGE and transferred to a nitrocellulose membrane by semidry blotting. On the membrane, the band corresponding to asFP595 appears red under daylight conditions and exhibits orange fluorescence with an emission maximum at 595 nm under UV light (Fig. 4 A and C). The fluorescence is stable and can even be detected after passage of the renatured protein through the membrane because of extended blotting. Renaturation with full restoration of fluorescence in the presence of β-mercaptoethanol has been described for heat-denatured GFP (31). Surprisingly, asCP562, which is nonfluorescent in vivo, behaves in the same manner and shows identical fluorescence as renatured asFP595 (Fig. 4 A and C). 31. Surpin, M. A. & Ward, W. W. (1989) Photochem. Photobiol. 49, 65. |
The source is mentioned three times, but it is not clear to the reader that the entire page is taken from it. |
| [10.] Tim/Fragment 011 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 11, Zeilen: 1-28 |
Quelle: Wiedenmann 2000 Seite(n): 14095, Zeilen: l.col: 1 ff. |
|---|---|
| The authors (Wiedenmann et al., 2000) noted that the two green fluorescent pigments can be distinguished by their spectral properties in the tentacles of A. sulcata var. rufescens although they could not be separated. This indicates that the two proteins have identical molecular weights or that they reflect two states of the same protein. The second hypothesis is supported by the construction of a cDNA library. In fact they assumed the existence of a second stable conformation of AsGFP499, with red-shifted fluorescence, which can be formed only in the expressing cells of the sea anemone.
Also the orange fluorescent protein asFP595 and the no [sic] fluorescent red protein asCP562 most likely represent two states of the same protein. Both proteins have the same molecular weight. A striking argument for their relationship is the finding that after a denaturation-renaturation process both asFP595 and asCP562 exhibit orange fluorescence with an emission maximum that matches that of asFP595. Therefore, the primary structure of asCP562 also must carry all features necessary for orange fluorescence. The putative semi-β-can structure of asCP562 can give a possible explanation of these phenomena. For GFP fluorescence the β-can structure is essential. They proposed the formation of a β-can-like structure in a multimerization process. This type of β-can consists of at least two molecules of asCP562. Their hypothesis was supported by the finding that both asCP562 and asFP595 show apparent molecular masses of 66 kDa. This molecular mass corresponds exactly to a tetramer consisting of four molecules asCP562 (4*16.5 kDa) (Table 1). Under the conditions necessary for renaturation of asFP595 a second fluorescent band can be observed if the samples are denatured without the presence of β-mercaptoethanol (fig. 4B). The second band migrates at twice the molecular mass of asFP595. This result confirms the hypothesis that asFP595 consists of at least two asCP562 monomers. Unlike GFP dimers, in asFP595 disulfide bonds seem to be involved in dimer interactions as a complete splitting is only possible under reducing conditions. However, to obtain the fluorescent state, it seems likely that a β-can-like structure is formed in which at least two molecules of asCP562 are linked with a disulfide bond. |
Two green fluorescent pigments can be distinguished by their spectral properties in the tentacles of A. sulcata var. rufescens although they could not be separated (Fig. 2A). This indicates that the two proteins have identical molecular weights or that they reflect two states of the same protein. The second hypothesis is supported by the construction of a cDNA library. It resulted in the cloning of several hundred GFPs of the asFP499 type. As the content of asFP499 and asFP522 in the tissue of tentacles is comparably high, one would expect that this ratio should be detectable in the library as well. We therefore assume the existence of a second stable conformation of asFP499, with red-shifted fluorescence, which can be formed only in the expressing cells of the sea anemone.
Also the orange fluorescent protein asFP595 and the non-fluorescent red protein asCP562 most likely represent two states of the same protein. Both proteins have the same molecular weight. A striking argument for their relationship is the finding that after a denaturation-renaturation process both asFP595 and asCP562 exhibit orange fluorescence with an emission maximum that matches that of asFP595. Therefore, the primary structure of asCP562 also must carry all features necessary for orange fluorescence. The putative semi-β-can structure of asCP562 can give a possible explanation of these phenomena. For GFP fluorescence the β-can structure is essential. [...] We propose the formation of a β-can-like structure in a multimerization process. This type of β-can consists of at least two molecules of asCP562. Together they could form a β-can with at least 12 β-strands surrounding at least two stretches homologous to the fluorophore region of GFP. Our hypothesis is supported by the finding that both asCP562 and asFP595 show apparent molecular masses of 66 kDa (Fig. 3.1 and 3.2). This molecular mass corresponds exactly to a tetramer consisting of four molecules asCP562 (4 x 16.5 kDa) (Table 1). Under the conditions necessary for renaturation of asFP595 a second fluorescent band can be observed if the samples are denatured without the presence of β-mercaptoethanol (Fig. 4B) The second band migrates at twice the molecular mass of asFP595. This result confirms the hypothesis that asFP595 consists of at least two asCP562 monomers. Unlike GFP dimers, in asFP595 disulfide bonds seem to be involved in dimer interactions as a complete splitting is only possible under reducing conditions. [...] However, to obtain the fluorescent state, it seems likely that a β-can-like structure is formed in which at least two molecules of asCP562 are linked with a disulfide bond. |
The source is given at the beginning of the page, but without clear indication that is meant to reference any more than maybe the first paragraph of the page. |
| [11.] Tim/Fragment 012 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 12, Zeilen: 1-2, 3-15 |
Quelle: Nienhaus 2006 Seite(n): 4213, 4215, 4216, Zeilen: 4213: r.col: 17 ff.; 4215: r.col.: 8 ff.; 4216: l.col. 1 ff. |
|---|---|
| [At pH ˃ 10, a red-]shifted [sic] emerged, with absorbance and emission peaks displaced by 18 and 10 nm to the red, respectively. At pH ˂ 4 and pH ˃ 12 the protein was thermodynamically unstable.
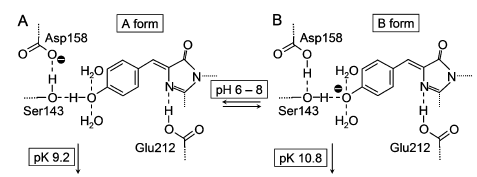
[...] In analogy to wild-type AvGFP, the AsGFP499 protein showed two bands in the UV/visible spectrum over a wide pH range. Although initially debated (Voityuk et al., 1998), there is now general agreement that the A and B bands are associated with the neutral and anionic states of chromophore (Tsien, 1998). In AsGFP499, the two bands were similar in area in the entire pH range in which the protein was stable; and remarkably, the protonated form of the chromophore became more dominant with increasing pH. In the structure of AsGFP499, amino acid Glu212 is shown to be hydrogen bonded to the chromophore heterocycle and not connected to the phenolic oxygen at all. Therefore, proton transfer between the chromophore and Glu212 cannot take place. In addition to hydrogen bonds to two water molecules, the Tyr64 phenol oxygen is connected to the Ser143 hydroxyl via a short hydrogen bond, which in turn is hydrogen bonded to Asp158. |
At pH . 10, a red-shifted form emerges, with absorbance and emission peaks displaced by 18 and 10 nm to the red, respectively (Fig. 2 D).
[page 4215] In analogy to wild-type avGFP, the asFP499 protein shows two bands in the UV/visible spectrum over a wide pH range (Fig. 4). Although initially debated (51,52), there is now general agreement that the A and B bands are associated with the neutral and anionic states of chromophore (9). [...] [...] In asFP499, the two bands are similar in area in the entire pH range in which the protein is stable (Fig. 4); and remarkably, the protonated form of the chromophore becomes more dominant with increasing pH. In the structure of asFP499, amino acid Glu212 (corresponding to Glu222 in avGFP) is shown to be hydrogen bonded to the chromophore heterocycle and not connected to the phenolic oxygen at all. Therefore, proton transfer between the chromophore and Glu212 cannot take place. However, the structure in Fig. 1 C suggests an alternative explanation for the appearance of the two conformations. In addition to hydrogen bonds to two water molecules, the Tyr64 phenol oxygen is connected to the [page 4216] Ser143 hydroxyl via a short hydrogen bond (2.5 A° ), which in turn is hydrogen bonded to Asp158 (2.7 A° ). 9. Tsien, R. Y. 1998. The green fluorescent protein. Annu. Rev. Biochem. 67:509–544. 51. Weber, W., V. Helms, J. A. McCammon, and P. W. Langhoff. 1999. Shedding light on the dark and weakly fluorescent states of green fluorescent proteins. Proc. Natl. Acad. Sci. USA. 96:6177–6182. 52. Voityuk, A. A., M. E. Michel-Beyerle, and N. Ro¨sch. 1998. Quantum chemical modeling of structure and absorption spectra of the chromophore in green fluorescent proteins. Chem. Phys. 231:13–25. |
The source is given on the previous page. The reader may assume that the work of Nienhaus et al. (2006) is described in the first two paragraphs, but not that this is done in the words of Nienhaus et al. (2006). The last documented paragraph however is not written in the descriptive past tense but rather makes factual statements, such that the reader has no way of knowing that he is reading the words of Nienhaus et al. (2006). |
| [12.] Tim/Fragment 013 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 13, Zeilen: 1-10 |
Quelle: Nienhaus 2006 Seite(n): 4216, Zeilen: l.col.: 2 ff. |
|---|---|
| The schemes in figure 5, A and B, show the tightly coupled system of two protonatable groups between which protons can be shuttled. The small ratio between neutral and anionic population implies that only slight differences in free energies exist between the two conformations in the electronic ground state.
Fig. 5. Schematic representation of the different protonation states of the AsGFP499 chromophore and its environment that are proposed to cause the spectral changes. (Nienhaus et al., 2006). Upon photon absorption, this balance is disturbed. Phenols typically become more acidic upon electronic excitation (Tsien, 1998; Voityuk et al., 1998); and therefore, we expect efficient excited state proton transfer (ESPT) to Asp158, as is inferred from the observation that excitation in the A and B bands is equally efficient for fluorescence in the 499nm emission band for pH ˂ 8. Evidently, protonation of Asp158 is a key ingredient in the proton shuttling mechanism described above. |
The schemes in Fig. 6, A and B, show the tightly coupled system of two protonatable groups between which protons can be shuttled. The small ratio between neutral and anionic population implies that only slight differences in free energies exist between the two conformations in the electronic ground state. Upon photon absorption, this balance is disturbed. Phenols typically become more acidic upon electronic excitation (9,52); and therefore, we expect efficient excited state proton transfer (ESPT) to Asp158, as is inferred from the observation that excitation in the A and B bands is equally efficient for fluorescence in the 499-nm emission band for pH < 8.
To further support the model presented in Fig. 6 by experimental evidence, we have produced the mutant Asp158Asn, which has its protonatable carboxyl residue replaced by a nonprotonatable carboxamide. Evidently, protonation of Asp158 is a key ingredient in the proton shuttling mechanism described above.
FIGURE 6 Schematic representation of the different protonation states of the asFP499 chromophore and its environment that are proposed to cause the spectral changes in Fig. 4. 9. Tsien, R. Y. 1998. The green fluorescent protein. Annu. Rev. Biochem. 67:509–544. 52. Voityuk, A. A., M. E. Michel-Beyerle, and N. Ro¨sch. 1998. Quantum chemical modeling of structure and absorption spectra of the chromophore in green fluorescent proteins. Chem. Phys. 231:13–25. |

The source is given for the figure and its caption, but not for the remainder of the text. |
| [13.] Tim/Fragment 013 12 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 13, Zeilen: 12-15 |
Quelle: Andresen et al 2005 Seite(n): 13070, Zeilen: l.col: 27 ff. |
|---|---|
| Recently, novel GFP-like fluorescent proteins have been discovered (Chudakov et al., 2003; Lukyanov et al. 2000; Ando et al., 2004) that can be reversibly photoswitched between a fluorescent (on) and nonfluorescent (off) state, that is, they are optically bistable and fluorescent. | Recently, novel GFP-like fluorescent proteins have been discovered (4–6) that can be reversibly photoswitched between a fluorescent (on) and nonfluorescent (off) state, that is, they are optically bistable and fluorescent.
4. Ando, R., Mizuno, H. & Miyawaki, A. (2004) Science 306, 1370–1373. 5. Chudakov, D. M., Belousov, V. V., Zaraisky, A. G., Novoselov, V. V., Staroverov, D. B., Zorov, D. B., Lukyanov, S. & Lukyanov, K. A. (2003) Nat. Biotechnol. 21, 191–194. 6. Lukyanov, K. A., Fradkov, A. F., Gurskaya, N. G., Matz, M. V., Labas, Y. A., Savitsky, A. P., Markelov, M. L., Zaraisky, A. G., Zhao, X. N., Fang, Y., et al. (2000) J. Biol. Chem. 275, 25879–25882. |
The source is given at the end of the next paragraph on the next page. |
| [14.] Tim/Fragment 014 05 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 14, Zeilen: 5-9 |
Quelle: Lukyanov 2000 Seite(n): 25880, Zeilen: l.col.: 21 ff. |
|---|---|
| With the exception of a slight shoulder at 530 nm, the protein displays a single absorption wavelength maximum, which occurs at 572 nm (Ԑ572 = 56,200 M-1 cm-1) (fig. 6). As is evident asFP595 absorbs efficiently in the middle range of the visible spectrum but remains translucent when excited with blue or long wavelength red light. Consequently, to the observer, the protein appears intensely purple.
[asFP595 can be transferred by green light from a nonfluorescent off into a fluorescent on state from which it reverts back eventually, but this transition can also be promptly stimulated by gentle irradiation with blue light (Lukyanov et al., 2000).] |
With the exception of a slight shoulder at 530 nm, the protein displays a single absorption wavelength maximum, which occurs at 572 nm (e572 = 56,200 M-1 cm-1). As is evident from this analysis, asFP595 absorbs efficiently in the middle range of the visible spectrum but remains translucent when excited with blue or long wavelength red light. Consequently, to the observer, the protein appears intensely purple. |
The source is mentioned at the end of the next sentence in the next paragraph, which, however, has been taken from another source: Tim/Fragment 014 10 |
| [15.] Tim/Fragment 014 10 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 14, Zeilen: 10-15 |
Quelle: Andresen et al 2005 Seite(n): 13070, Zeilen: 44 ff. |
|---|---|
| asFP595 can be transferred by green light from a nonfluorescent off into a fluorescent on state from which it reverts back eventually, but this transition can also be promptly stimulated by gentle irradiation with blue light (Lukyanov et al., 2000). The "on-off" cycle can be repeated many times. However, with its low quantum yield (˂0.001), and comparatively slow switching kinetics, the photochromic properties of asFP595 are far from being optimal. | It can be transferred by green light from a nonfluorescent off into a fluorescent on state from which it reverts back eventually, but this transition can also be promptly stimulated by gentle irradiation with blue light (6). The "on-off" cycle can be repeated many times. However, with its low quantum yield (<0.001, ref. 6) and comparatively slow switching kinetics, the photochromic properties of asFP595 are far from being optimal.
6. Lukyanov, K. A., Fradkov, A. F., Gurskaya, N. G., Matz, M. V., Labas, Y. A., Savitsky, A. P., Markelov, M. L., Zaraisky, A. G., Zhao, X. N., Fang, Y., et al. (2000) J. Biol. Chem. 275, 25879–25882. |
The source is only mentioned in the next paragraph. |
| [16.] Tim/Fragment 014 19 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 14, Zeilen: 19-24, 27-33 |
Quelle: Andresen et al 2005 Seite(n): 13071, Zeilen: r.col. 14 ff. |
|---|---|
| The quantum yield of these variants was major as compared with that of the wt asFP595 protein. As in all GFP-like proteins, the asFP595 chromophore resides in a helical segment, which is enclosed in an 11-stranded β-barrel (Ormo et al., 1996; Wall et al., 2000; Yang et al., 1996). The tripeptide M63-Y64-G65 rearranges to a chromophoric, conjugated 2-iminomethyl-5-(4-hydroxybenzylidene) imidazolinone (MYG) system.
[...] Unlike most other GFP-like proteins, the protein backbone is broken between C62 and the chromophore. Nevertheless, the former M63 Cα and backbone nitrogen atoms are in plane with the imidazolinone ring and, thus, part of the conjugated system. The imino group expands the conjugated system of MYG, likely accounting for the shift of the absorption maximum toward a longer wavelength (572 nm) as compared with that of GFP (470 nm). The MYG chromophore of the off state asFP595 exclusively adopts the trans conformation. |
As in all GFP-like proteins, the asFP595 chromophore resides in a helical segment, which is enclosed in an 11-stranded β-barrel (20–23). The tripeptide M63-Y64-G65 rearranges to a chromophoric, conjugated 2-iminomethyl-5-(4-hydroxybenzylidene) imidazolinone (MYG) system, in agreement with previous biochemical analysis of proteolytic peptides (24). Unlike most other GFP-like proteins, the protein backbone is broken between C62 and the chromophore. Nevertheless, the former M63 Cα and backbone nitrogen atoms are in plane with the imidazolinone ring and, thus, part of the conjugated system. The imino group expands the conjugated system of MYG, likely accounting for the shift of the absorption maximum toward a longer wavelength (572 nm) as compared with that of GFP (470 nm). The MYG chromophore of the off state asFP595 exclusively adopts the trans conformation.
[...] [...] This variant has 12 times the fluorescence quantum yield as compared with that of the wt asFP595 protein (6). 6. Lukyanov, K. A., Fradkov, A. F., Gurskaya, N. G., Matz, M. V., Labas, Y. A., Savitsky, A. P., Markelov, M. L., Zaraisky, A. G., Zhao, X. N., Fang, Y., et al. (2000) J. Biol. Chem. 275, 25879–25882. 20. Ormo, M., Cubitt, A. B., Kallio, K., Gross, L. A., Tsien, R. Y. & Remington, S. J. (1996) Science 273, 1392–1395. 21. Wall, M. A., Socolich, M. & Ranganathan, R. (2000) Nat. Struct. Biol. 7, 1133–1138. 22. Yang, F., Moss, L. G. & Phillips, G. N., Jr. (1996) Nat. Biotechnol. 14, 1246–1251. 23. Yarbrough, D., Wachter, R. M., Kallio, K., Matz, M. V. & Remington, S. J. (2001) Proc. Natl. Acad. Sci. USA. 98, 462–467. 24. Zagranichny, V. E., Rudenko, N. V., Gorokhovatsky, A. Y., Zakharov, M. V., Balashova, T. A. & Arseniev, A. S. (2004) Biochemistry 43, 13598– 13603. |
The source is mentioned right before the documented passage. One could assume that the first documented sentence describes the results of Andresen et al. (2005), what follows, however, is attributed to other sources. |
| [17.] Tim/Fragment 015 11 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 15, Zeilen: 11-15 |
Quelle: Schaefer 2007 Seite(n): 540, Zeilen: l.col.: 3 ff. |
|---|---|
| The proposed kindling mechanism (Schafer et al., 2007; Andresen et al., 2005) is depicted in figure 8. Before the photon absorption the chromophore is in the dark zwitterionic trans state but when the photon arrives it changes in the cis state. The isomerization increases the acidity of the imidazolinone NH proton of the zwitterionic cis chromophore and induces proton transfer to Glu215 (state A-cis, figure 8).
Whether this proton transfer takes place in the electronic ground state or in the excited state cannot be judged on the basis of the current simulations. The proton transfer also [prevents photoisomerization of the zwitterionic cis species back to the initial dark trans state.] |
The proposed kindling mechanism is depicted in Figure 3. As shown previously, kindling is initiated by a trans–cis photoisomerization through a space-saving hula-twist mechanism.[ 11] The isomerization increases the acidity of the imidazolinone NH proton of the zwitterionic cis chromophore and induces proton transfer to Glu215 (state A-cis, Figure 3). Whether this proton transfer takes place in the electronic ground state or in the excited state cannot be judged on the basis of the current simulations. The proton transfer also prevents photoisomerization of the zwitterionic cis species back to the initial dark trans state. |
The source is given together with another source such that a literal quote cannot be understood. This is particularly the case in the second paragraph, as it describes "current simulations". |
| [18.] Tim/Fragment 016 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 16, Zeilen: 1-7 |
Quelle: Schaefer 2007 Seite(n): 540, Zeilen: l.col.: 11 ff. |
|---|---|
| [The proton transfer also] prevents photoisomerization of the zwitterionic cis species back to the initial dark trans state. The anionic cis intermediate is subsequently protonated at the phenolate oxygen atom to finally yield the stable neutral chromophore (state N-cis).
This situation is similar to that in GFP, where the neutral protonation state is also favored for the cis chromophore (Helms, 2002; Stoner-Ma et al., 2005; Agmon, 2005; Leiderman et al., 2006; Vendrell et al., 2006). GFP fluorescence originates from the anionic chromophore, which is formed through excited-state proton transfer (ESPT). |
The proton transfer also prevents photoisomerization of the zwitterionic cis species back to the initial dark trans state. The anionic cis intermediate is subsequently protonated at the phenolate oxygen atom to finally yield the stable neutral chromophore (state Ncis). This situation is similar to that in GFP, where the neutral protonation state is also favored for the cis chromophore.[16–20] GFP fluorescence originates from the anionic chromophore, which is formed through excited-state proton transfer (ESPT).
[16] V. Helms, Curr. Opin. Struct. Biol. 2002, 12, 169. [17] D. Stoner-Ma, A. A. Jaye, P. Matousek, M. Towrie, S. R. Meech, P. J. Tonge, J. Am. Chem. Soc. 2005, 127, 2864. [18] N. Agmon, Biophys. J. 2005, 88, 2452. [19] P. Leiderman, D. Huppert, N. Agmon, Biophys. J. 2006, 90, 1009. [20] O. Vendrell, R. Gelabert, M. Moreno, J. M. Lluch, J. Am. Chem. Soc. 2006, 128, 3564. |
The source is given on the previous page. |
| [19.] Tim/Fragment 017 15 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 17, Zeilen: 15-16 |
Quelle: Isarankura 2009 Seite(n): 353, Zeilen: 17 ff. |
|---|---|
| Recently, the fluorescent proteins from various organisms become potential candidates for sensor development. These [include the green fluorescent protein and its variants, the red fluorescent protein (DsRed) from the tropical coral namely Discosoma sp.] | Recently, the fluorescent proteins from various organisms become potential candidates for sensor development. These include the green fluorescent protein (GFP) and its variants from the Pacific Northwest jellyfish namely Aequorea victoria [7–11], the red fluorescent protein (DsRed) from the tropical coral namely Discosoma sp. [12–14], and the far-red fluorescent (HcRed) protein from the reef coral namely Heteractis crispa [15, 16]. |
The source is not given here, but will be mentioned on page 19. The text of the source has been shortened in a way that is little consistent with normal English language usage. |
| [20.] Tim/Fragment 019 01 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 19, Zeilen: 1-2 |
Quelle: Chapleau 2008 Seite(n): 619, Zeilen: l.col.: 14 ff. |
|---|---|
| Previous studies have suggested that the irregular conformation of the structure might allow solutes to penetrate into the core of the protein (Agmon, 2005). | Previous studies have suggested that the irregular conformation of the structure might allow solutes to penetrate into the core of the protein (Agmon 2005). |
|
| [21.] Tim/Fragment 019 03 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 19, Zeilen: 3-16 |
Quelle: Isarankura 2009 Seite(n): 352, Zeilen: abstract |
|---|---|
| Isarankura et al. (2009) explored spectroscopic determinations of copper ions using chimeric metal-binding green fluorescent protein His6GFP (espressed in E. coli and purified to homogeneity) as an active indicator. Supplementation of copper ions to the GFP solution led to a remarkable decrease of fluorescent intensity corresponding to metal concentrations. For circumstances, rapid declining of fluorescence up to 60% was detected in the presence of 500 μM copper. This was in contrast to those observed in the case of zinc and calcium ions, in which approximately 10–20% of fluorescence was affected (fig. 11). Recovery of its original fluorescence up to 80% was mediated by the addition of EDTA.
More importantly, in the presence of metal ions, the emission wavelength maximum remained unchanged while reduction of the optical density of the absorption spectrum has been observed. This indicated that the chromophore‟s ground state was possibly affected by the static quenching process rather than structural or conformational alteration. |
Abstract
In the present study, spectroscopic determinations of copper ions using chimeric metal-binding green fluorescent protein (His6GFP) as an active indicator have been explored. Supplementation of copper ions to the GFP solution led to a remarkable decrease of fluorescent intensity corresponding to metal concentrations. For circumstances, rapid declining of fluorescence up to 60% was detected in the presence of 500 μM copper. This is in contrast to those observed in the case of zinc and calcium ions, in which approximately 10–20% of fluorescence was affected. Recovery of its original fluorescence up to 80% was mediated by the addition of ethylenediamine tetraacetic acid. More importantly, in the presence of metal ions, the emission wavelength maximum remains unchanged while reduction of the optical density of the absorption spectrum has been observed. This indicates that the chromophore’s ground state was possibly affected by the static quenching process. |
The source is given, but it is not clear to the reader that the following text is taken from it. The copied text includes, what could be understood as an interpretation / evaluation of the results of the source: "a remarkable decrease", "More importantly", "This indicated that", "possibly". |
| [22.] Tim/Fragment 020 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 20, Zeilen: 1-15 |
Quelle: Sumner et al 2006 Seite(n): 1303, 1306, Zeilen: 1303: l.col: 25 ff.; 1305: l.col: 21 ff.; 1306: r.col: 7 ff. |
|---|---|
| Sumner et al. (2006) described the quenching of the fluorescence of DsRed (Fradkov et al., 2002) by Cu+ and Cu2+. They showed that the DsRed tetramer can be used as a sensitive fluorescent probe towards both monovalent and divalent copper ions, even in the presence of competing metal ions, with a detection limit well below 1 ppb. The wild type DsRed is 40 times more sensitive than the engineered GFPs and seven orders of magnitude more sensitive to Cu2+ ions than the wild type GFP. Even more impressive is the ability of DsRed to bind Cu+ with nanomolar sensitivity and a fast response time. They for first demonstrated that the addiction of Mn2+, Fe2+/3+, Co2+, Ni2+, Zn2+, Cd2+, Ag+, Hg2+, Pb2+, Mg2+ or Ca2+ did not competed for the copper binding site and did not interfered under these conditions. These data also indicated that the quenching was not a result of collisional quenching. With EDTA the fluorescence intensity returned to 90% of its initial value. Copper ions prefer coordination with amines, ionized peptide nitrogens and thiolates (Silvia and Williams, 1991); structural motifs that have high affinity for Cu2+ are frequently composed of the side chains of His, Tyr, Glu, Asp, Cys, and sometimes Asn and Gln (Sigel and Martin, 1982). DsRed contains a single cysteine [which is solvent accessible and many solvent accessible His, Glu, and Asp, as shown by the X-ray structure (Wall et al., 2000).] | In this report, we describe the quenching of the intrinsic fluorescence of DsRed (Fradkov et al., 2002) by Cu+ and Cu2+; [...] Here we show that the DsRed tetramer can be used as a sensitive fluorescent probe towards both monovalent and divalent copper ions, even in the presence of competing metal ions, with a detection limit well below 1 ppb. The wild type DsRed is 40 times more sensitive than the engineered GFPs and seven orders of magnitude more sensitive to Cu2+ ions than the wild type GFP. Even more impressive is the ability of DsRed to bind Cu+ with nanomolar sensitivity and a fast response time.
[page 1305] As an initial test of the selectivity, we first demonstrated that the DsRed fluorescence is unaffected by the addition of 1μM of Mn2+, Fe2+/3+, Co2+, Ni2+, Zn2+, Cd2+, Ag+, Hg2+, Pb2+, or 1μM Mg2+ or Ca2+. [...] Furthermore, other transition metals do not affect the Cu2+-mediated quenching of DsRed (Fig. 3, front row), indicating that these metals do not compete for the copper binding site and do not interfere under these conditions. [...] [...] The fluorescence intensity, after being quenched by copper(II), returns to 90% of its initial value within 2 min when EDTA, a metal ion chelator, is added and allowed to equilibrate. [page 1306] Copper ions prefer coordination with amines, ionized peptide nitrogens and thiolates (Silvia and Williams, 1991); structural motifs that have high affinity for Cu2+ are frequently composed of the side chains of His, Tyr, Glu, Asp, Cys, and sometimes Asn and Gln (Sigel and Martin, 1982) and yield square planar, trigonal bipyramid and octahedral geometries. DsRed contains a single cysteine which is solvent accessible and many solvent accessible His, Glu, and Asp, as shown by the X-ray structure (Wall et al., 2000). |
The source is mentioned in the beginning, but without making clear that the whole page is taken from it largely literally. In the beginning of the fragment the work of Sumner et al. (2006) is described and the reader may expect a certain closeness, but starting with "Copper ions prefer" findings of other papers are described and the reader cannot assume in any way that this passage is also a literal copy of Sumner et al. (2006). |
| [23.] Tim/Fragment 023 03 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 23, Zeilen: 3-5 |
Quelle: Isarankura 2009 Seite(n): 353, Zeilen: last lines |
|---|---|
| Engineered E. coli co-expressing surface exposed zinc-binding motif and GFP has been applied for real-time monitoring of intracellular mobility of zinc ions (Isarankura-Na-Ayudhya et al., 2005).
Isarankura-Na-Ayudhya C., Suwanwong Y., Boonpangrak S., Kiatfuengfoo R., Prachayasittikul V., 2005. Co-expression of zinc binding motif and GFP as a cellular indicator of metal ions mobility. Int. J. Biol. Sci., 1: 146-151. |
Additionally, engineered E. coli co-expressing surface exposed zinc-binding motif and GFP has been applied for real-time monitoring of intracellular mobility of zinc ions [27].
27. Isarankura-Na-Ayudhya C, Suwanwong Y, Boonpangrak S et al (2005) Co-expression of zinc binding motif and GFP as a cellular indicator of metal ions mobility. Int J Biol Sci 1:146–151 |
The source given does not contain the copied text. |
| [24.] Tim/Fragment 023 19 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 23, Zeilen: 19-26 |
Quelle: Fu 2008 Seite(n): abstract, p.6, Zeilen: p.6: last lines |
|---|---|
| In the study of 2008, Fu et al. observed the proximity of single GFPs to metallic silver nanoparticles increases its fluorescence intensity approximately 6-fold and decreases the decay time. Single protein molecules on the silvered surfaces emitted 10-fold more photons as compared to glass prior to photobleaching. The photostability of single GFP has increased to some extent. Accordingly, we observed longer duration time and suppressed blinking. The detection of metal-surface enhanced fluorescence from GFP suggests the more extensive use of metallic nanostructures in imaging and single molecule detection. | In this report, we observed the proximity of single GFPs to metallic silver nanoparticles increases its fluorescence intensity approximately 6-fold and decreases the decay time. Single protein molecules on the silvered surfaces emitted 10-fold more photons as compared to glass prior to photobleaching. The photostability of single GFP has increased to some extent. Accordingly, we observed longer duration time and suppressed blinking.
The detection of metal-surface enhanced fluorescence from GFP suggests the more extensive use of metallic nanostructures in imaging and single molecule detection. |
The source is given, but it is not clear to the reader how much has been taken from it. In particular the two sentences starting with "Accordingly, we observed" cannot be attributed to the source by the reader who rather assumes that the author herself made observations that are consistent with the findings of the source. |
| [25.] Tim/Fragment 024 06 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 24, Zeilen: 6-11 |
Quelle: MSIP 2010 Seite(n): 1 (online source), Zeilen: - |
|---|---|
| It has about 100 radial canals and about 3-6 times as many tentacles that are bulb laterally compressed. Umbrella up to 145 mm wide is usually 60-80 mm.
[...] The low and thick manubrium is about half as wide as umbrella. The gonads extend along almost whole length of radial canals. It has numerous small bulbs with excretory papillae and statocysts numerous, crowded. |
About 100 radial canals, 3-6 times as many tentacles; tentacle bulbs laterally compressed. Umbrella up to 145 mm wide, usually 60-80 mm, low and thick manubrium, about half as wide as umbrella; gonads extending along almost whole length of radial canals; numerous small bulbs; all bulbs with excretory papillae; statocysts numerous, crowded. |
The source is not mentioned. |
| [26.] Tim/Fragment 024 09 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 24, Zeilen: image |
Quelle: Wikipedia Conica (suborder) 2009 Seite(n): 1 (online source), Zeilen: - |
|---|---|

Fig. 13. Aequorea coerulescens. |

Aequorea coerulescens medusa (Aequoreidae) |
The source of the image is not given. The image is available under the Attribution-ShareAlike 3.0 Licence, which allows free usage but demands that appropriate credit is given. |
| [27.] Tim/Fragment 025 01 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 25, Zeilen: 1 ff. (entire page) |
Quelle: Gurskaya 2003 Seite(n): 404, Zeilen: l.col. last lines |
|---|---|
| [Although A. coerulecens and A. victoria look similar, some of their] features are clearly different. The most obvious difference is that A. victoria carries only 1 tentacle in the radial channel, whereas A. coerulescens possesses 3–6 tentacles between each pair of adjacent radial channels.
In contrast to A. victoria, A. coerulescens medusae displayed blue, not green, luminescence. Gurskaya et al. (2003) described for the wild type acGFP no detectable fluorescence under either UV light or using a fluorescence microscope. However, using monoclonal antibodies against GFP they found that a protein extract from A. coerulescens contained a GFP-like protein that had the 92% identity at the amino acid level with AvGFP. It was detected in the umbrella border. All known key GFP residues are conserved in AcGFP, including the chromophore forming Ser65, Tyr66 and Gly67 residues, the evolutionarily invariant Arg96 and Glu222, and His148, Phe165, Ile167 and Thr203 which are all spatially close to the chromophore. Gurskaya et al. (2003) used a random mutagenesis to generate fluorescent mutants of wt GFP of this species. The substituting E222G appeared to be the key event in creating a fluorescent form. Moreover five amino-acid substitutions compared with the wild-type, specifically V11I, F64L, K101E, T206A and E222G determined a very bright mutant. |
Although A. coerulecens and A. victoria (other names for the latter are A. forscalea and A. aequorea) look similar, some of their features are clearly different. The most obvious difference is that A. victoria carries only 1 tentacle per radial channel, whereas A. coerulescens possesses 4–6 tentacles between each pair of adjacent radial channels [15].
In contrast to A. victoria, A. coerulescens medusae displayed blue, not green, luminescence. We cloned the aequorin-like photoprotein apparently responsible for the bioluminescence observed in A. coerulescens specimens (GenBank® accession number AY236998). This molecule shared 84% identity at the amino acid level with A. victoria aequorin, and had an emission maximum at 455 nm. No detectable fluorescence was observed in A. coerulescens under either UV light or using a fluorescence microscope. However, using monoclonal antibodies against GFP we found that a protein extract from A. coerulescens contained a GFP-like protein (see Figure 3A). [...] [...] All known key GFP residues are conserved in acGFPL, including the chromophore-forming Ser65, Tyr66 and Gly67 residues, the evolutionarily invariant Arg96 and Glu222, and His148, Phe165, Ile167 and Thr203 which are all spatially close to the chromophore. [...] [...] Random mutagenesis was used to improve protein folding and to generate fluorescent mutants of acGFPL. After the first round of mutagenesis several green fluorescent colonies of different brightness were found. The three brightest clones (named G1, Z1 and Z2) were further characterized and the mutants were found to contain the same substitution, E222G. Additional V11I/K101E and N19D mutations were found in G1 and Z2 respectively. Thus substituting Glu222 appeared to be the key event in creating a fluorescent form of acGFPL. [...] Two additional rounds of random mutagenesis on the G1 clone (only G1 was able to mature at 37 ◦C, whereas Z1 and Z2 required incubation at low temperature to develop fluorescence) generated a very bright mutant, named aceGFP, which matured fast at 37 ◦C. This mutant contained five amino-acid substitutions compared with the wild-type acGFPL, specifically V11I, F64L, K101E, T206A and E222G (Figure 1). 15 Kramp, P. L. (1968) The hydromedusae of the Pacific and Indian Oceans. Dana Rept. 72, 201–202 |
The source is given and from the context it becomes clear that some results of Gurskaya et al. (2003) are reported. However, it is not clear that this reporting is done using the words of Gurskaya et al. (2003) and there are also passages taken from Gurskaya et al. (2003) that the reader would not attribute to Gurskaya et al. (2003) (first and third paragraph). |
| [28.] Tim/Fragment 041 08 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 41, Zeilen: 8-10 |
Quelle: Wikipedia Snakelocks anemone 2012 Seite(n): online source, Zeilen: - |
|---|---|
| Its retractile tentacles are green colour with purple tips. This is due to the presence of symbiotic algae (zooxanthellae) within the tentacles that use sunlight as an energy source. Since the anemones benefit from this they prefer brightly lit shallow waters. | The tentacles of anemones in deep or murky water can be a grey colour but are otherwise usually a deep green colour with purple tips. This is due to the presence of symbiotic algae within the tentacles that use sunlight as an energy source. Since the anemones benefit from this they prefer brightly lit shallow waters. |
The source is not mentioned. Note that there are invisible, but active wikipedia links for "symbiosis", "algae" and "tentacles" in the PDF file of the thesis that is available for download. In the source one can find the same links. |
| [29.] Tim/Fragment 042 02 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 42, Zeilen: 2-18 |
Quelle: Wikipedia Snakelocks anemone 2012 Seite(n): online source, Zeilen: - |
|---|---|
| On average the anemone is 8 cm large.
Unlike other cnidarians, anemones (and other Anthozoa) entirely lack the free-swimming medusa stage of the life cycle; the polyp produces eggs and sperm, and the fertilized egg develops into a planula that develops directly into another polyp. Anemones tend to stay in the same spot until conditions become unsuitable (prolonged dryness, for example), or a predator attacks them. In that case, anemones can release themselves from the substrate and use flexing motions to swim to a new location. Most sea anemones attach temporarily to submerged objects; a few thrust themselves into the sand or live in furrows; a few are parasitic on other marine organisms. The sexes in sea anemones are separate in some species, while others are protandric hermaphrodites. The gonads are strips of tissue within the mesenteries. Both sexual and asexual reproduction can occur. In sexual reproduction males release sperm to stimulate females to release eggs, and fertilization occurs. Anemones eject eggs and sperm through the mouth. The fertilized egg develops into a planula, which settles and grows into a single polyp. In asexual reproduction extratentacular budding occurs, a type of fission, where the anemone will split roughly in half generally through the mouth. This species is extremely adaptable to various conditions of temperature and salinity. |
On average the snakelock anemone is 8 cm large.
[...] Unlike other cnidarians, anemones (and other Anthozoa) entirely lack the free-swimming medusa stage of the life cycle; the polyp produces eggs and sperm, and the fertilized egg develops into a planula that develops directly into another polyp. Anemones tend to stay in the same spot until conditions become unsuitable (prolonged dryness, for example), or a predator attacks them. In that case anemones can release themselves from the substrate and use flexing motions to swim to a new location. Most sea anemones attach temporarily to submerged objects; a few thrust themselves into the sand or live in furrows; a few are parasitic on other marine organisms.[1] The sexes in sea anemones are separate in some species, while others are protandric hermaphrodites. The gonads are strips of tissue within the mesenteries. Both sexual and asexual reproduction can occur. In sexual reproduction males release sperm to stimulate females to release eggs, and fertilization occurs. Anemones eject eggs and sperm through the mouth. The fertilized egg develops into a planula, which settles and grows into a single polyp. In asexual reproduction extratentacular budding occurs, a type of fission, where the anemone will split roughly in half generally through the mouth. [...] [...] It rapidly adapts to aquarium life as they are extremely adaptable to various conditions. |
The source is not mentioned. Note that there are invisible, but active wikipedia links for "cnidarians", "Anthozoa" and "medusa" in the PDF file of the thesis that is available for download. In the source one can find the same links. |
| [30.] Tim/Fragment 069 25 |
| Verschleierung |
|---|
| Untersuchte Arbeit: Seite: 69, Zeilen: 25-32 |
Quelle: Copper 2010 Seite(n): 1 (online source), Zeilen: - |
|---|---|
| Copper is one of the two exceptions to the writing of electron configurations. One would expect copper to be an s2d9 ion. Instead, one electron is borrowed from the 4s orbital to completely fill the 3d orbital. Copper metal is therefore paramagnetic due to the unpaired electron in the orbital [sic]. The two most common ions formed by copper are the +1 (cuprous) and +2 (cupric). The cuprous ion is the less stable of the two oxidation states and is easily oxidized. Since electrons are lost first out of the 4s orbital, the electron configuration of the cuprous ion is 4s03d10 and is therefore diamagnetic while the cupric ion is paramagnetic (4s03d9). | Copper is one of the two exceptions to the writing of electron configurations. One would expect copper to be an s2d9 ion. Instead, one electron is "borrowed" from the 4s orbital to completely fill the 3d orbitals. Copper metal is therefore paramagnetic due to the unpaired electron in the 4s orbital. [...]
[...] The two most common ions formed by copper are the +1 (cuprous) and +2 (cupric).The cuprous ion is the less stable of the two oxidation states and is easily oxidized. Therefore, any work done with copper(I) compounds must be carried out in an inert atmosphere. Since electrons are lost first out of the 4s orbital, the electron configuration of the cuprous ion is 4s03d10 and the [sic] it is therefore diamagnetic. [...] [...] Removal of a second electron results in the a 4s03d9 configuration. Therefore the cupric ion is paramagnetic. |
The source is not mentioned. |
| [31.] Tim/Fragment 070 29 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 70, Zeilen: 29-32 |
Quelle: Sumner et al 2006 Seite(n): 1306, Zeilen: r.col: 7 ff. |
|---|---|
| Copper ions prefer coordination with amines, ionized peptide nitrogens and thiolates (Silvia and Williams, 1991); structural motifs that have high affinity for Cu2+ are frequently composed of the side chains of His, Tyr, Glu, Asp, Cys, and sometimes Asn and Gln (Sigel and Martin, 1982). | Copper ions prefer coordination with amines, ionized peptide nitrogens and thiolates (Silvia and Williams, 1991); structural motifs that have high affinity for Cu2+ are frequently composed of the side chains of His, Tyr, Glu, Asp, Cys, and sometimes Asn and Gln (Sigel and Martin, 1982) |
The source is mentioned in the previous paragraph. The same passage can be found on page 20: Tim/Fragment 020 01 |
| [32.] Tim/Fragment 071 05 |
| BauernOpfer |
|---|
| Untersuchte Arbeit: Seite: 71, Zeilen: 5-14 |
Quelle: Rahimi et al 2008 Seite(n): 5, Zeilen: 15 ff. |
|---|---|
| It is known that in the case of proteins only the fluorophores located at the surface can be dynamically quenched whereas the internal fluorophores are quenched by static process due to higher hydrophobicity in the interior. In the case of AsGFP, the chromophore is well shielded inside a β-barrel and hence is not accessible for dynamic or collisional quenching. Rahimi et al. (2008) reported for DsRed, that Cu2+ forms a complex with specific amino acids on the protein. This complex formation can lead to formation of non-fluorescent species in three ways, i) by affecting hydrogen-bonding network of the chromophore, ii) by bringing Cu2+ close to chromophore such that it contacts the excited-state of the chromophore, iii) by causing conformation/structural changes in the protein. | It is known that in the case of proteins only the fluorophores located at the surface can be dynamically quenched whereas the internal fluorophores are quenched by static process due to higher hydrophobicity in the interior. In the case of DsRed, the chromophore is well shielded inside a β-barrel and hence is not accessible for dynamic or collisional quenching. [...] We hypothesize that, in DsRed, Cu2+ forms a complex with specific amino acids on the protein. This complex formation can lead to formation of non-fluorescent species in three ways, i) by affecting hydrogen-bonding network of the chromophore, ii) by bringing Cu2+ close to chromophore such that it contacts the excited-state of the chromophore, iii) by causing conformation/structural changes in the protein. |
The source is mentioned in the middle. The reader might assume that the text following the reference is close to the source, but would expect that the text before the reference is the view of the author. It is remarkable that Rahmini et al. (2008) "hypothesize that, in DsRed, Cu2+ forms a complex with specific amino acids on the protein", whereas T. M. writes: "Rahimi et al. (2008) reported for DsRed, that Cu2+ forms a complex with specific amino acids on the protein." From a scientific point of view, this is a significant misrepresentation. |
| [33.] Tim/Fragment 072 18 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 72, Zeilen: 18-32 |
Quelle: Shang Dong 2008 Seite(n): 1569, Zeilen: l.col, 1 ff. |
|---|---|
| There is an enormous demand for optical sensors in many areas, such as the environmental monitoring, industrial and food processing, healthcare, biomedical technology, and clinical analysis, where high sensitivity and ease of operation are two main issues (Shi et al., 2004; Jeronimo et al., 2007). Fluorescence technique can easily fulfill both requirements and has been widely applied in the fields of chemical sensing and biosensing (Jiang and Guo, 2004; Prodi, 2005; Lim and Lippard, 2007). In the design of fluorescence-based sensors, the fluorescence probes remain a key determinant to the future success of the sensor. Therefore, the development of fluorescence probes has been a long-term and attractive topic among the scientists (Callan et al., 2005; Demchenko, 2005). In this regard, many kinds of fluorophores have been developed, and already been applied in the sensors design. Unfortunately, the performance of these established fluorescent sensors are greatly limited by using the conventional fluorescence probes. For instance, organic fluorophores, which are most commonly used in fluorescence technique, suffer from low brightness and poor photostability, which lead to reduced sensitivity and stability of the sensor (Zhou and Zhou, 2004; [Demchenko, 2005; Prodi, 2005).] | There is an enormous demand for optical sensors in many areas, such as the environmental monitoring, industrial and food processing, healthcare, biomedical technology, and clinical analysis, where high sensitivity and ease of operation are two main issues (Shi et al., 2004; Jeronimo et al., 2007). Fluorescence technique can easily fulfill both requirements and has been widely applied in the fields of chemical sensing and biosensing (Jiang and Guo, 2004; Prodi, 2005; Lim and Lippard, 2007). In the design of fluorescence-based sensors, the fluorescence probes remain a key determinant to the future success of the sensor. Therefore, the development of fluorescence probes has been a long-term and attractive topic among the scientists (Callan et al., 2005; Demchenko, 2005). In this regard, many kinds of fluorophores have been developed, and already been applied in the sensors design. Unfortunately, the performance of these established fluorescent sensors are greatly limited by using the conventional fluorescence probes. For instance, organic fluorophores, which are most commonly used in fluorescence technique, suffer from low brightness and poor photostability, which lead to reduced sensitivity and stability of the sensor (Zhou and Zhou, 2004; Demchenko, 2005; Prodi, 2005). |
The source is not mentioned anywhere. |
| [34.] Tim/Fragment 073 01 |
| KomplettPlagiat |
|---|
| Untersuchte Arbeit: Seite: 73, Zeilen: 1 ff. (complete page) |
Quelle: Shang Dong 2008 Seite(n): 1569, Zeilen: l.col. 21 ff. |
|---|---|
| In addition, organic fluorophores tend to have small Stokes shifts, thus limiting their utility in the fields, such as ratiometric measurements and fluorescence resonance energy transfer (FRET)- based detection (Demchenko, 2005). Compared with conventional organic fluorophores, the recently developed semiconductor quantum dots show great promise as the fluorescence probes due to their improved photophysical properties, including high quantum yield, size-tunable narrow emissions and minimal photobleaching (Costa-Fernaˇındez et al., 2006; Somers et al., 2007). However, these nanocrystalline materials still suffer from several issues, such as the harsh and toxic conditions in the synthesis, large physical size, strong non-molecular low-power fluorescence intermittency, which also limit their further application in fluorescence sensing research (Verberk et al., 2002; Derfus et al., 2004; Vosch et al., 2007). Therefore, the development of new fluorophores that can challenge the above problems is of great importance for both fluorescence sensing and other fluorescence-based applications like fluorescence imaging (Rao et al., 2007). | In addition, organic fluorophores tend to have small Stokes shifts, thus limiting their utility in the fields, such as ratiometric measurements and fluorescence resonance energy transfer (FRET)- based detection (Demchenko, 2005). Compared with conventional organic fluorophores, the recently developed semiconductor quantum dots show great promise as the fluorescence probes due to their improved photophysical properties, including high quantum yield, size-tunable narrow emissions and minimal photobleaching (Costa-Fernaˇındez et al., 2006; Somers et al., 2007). However, these nanocrystalline materials still suffer from several issues, such as the harsh and toxic conditions in the synthesis, large physical size, strong non-molecular low-power fluorescence intermittency, which also limit their further application in fluorescence sensing research (Verberk et al., 2002; Derfus et al., 2004; Vosch et al., 2007). Therefore, the development of new fluorophores that can challenge the above problems is of great importance for both fluorescence sensing and other fluorescence-based applications like fluorescence imaging (Rao et al., 2007). |
The source is not mentioned anywhere. |
Sources
| [1.] Quelle:Tim/Shang Dong 2008 |
| Autor | Li Shang, Shaojun Dong |
| Titel | Sensitive detection of cysteine based on fluorescent silver clusters |
| Zeitschrift | Biosensors and Bioelectronics |
| Jahr | 2009 |
| Seiten | 1569–1573 |
| URL | http://www.ncbi.nlm.nih.gov/pubmed/18823770 |
Literaturverz. |
no |
| Fußnoten | no |
| [2.] Quelle:Tim/Wang 2007 |
| Autor | Wen Wang |
| Titel | Validation of shRNA clones for gene silencing in 293FT cells |
| Ort | Würzburg |
| Jahr | 2007 |
| URL | http://opus.bibliothek.uni-wuerzburg.de/frontdoor/index/index/docId/2225 |
Literaturverz. |
no |
| Fußnoten | no |
| [3.] Quelle:Tim/Nienhaus 2006 |
| Autor | Karin Nienhaus, Fabiana Renzi, Beatrice Vallone, Jörg Wiedenmann, and G. Ulrich Nienhaus |
| Titel | Chromophore-Protein Interactions in the Anthozoan Green Fluorescent Protein asFP499 |
| Zeitschrift | Biophysical Journal |
| Datum | December 2006 |
| Seiten | 4210–4220 |
| URL | http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1635683/pdf/4210.pdf |
Literaturverz. |
yes |
| Fußnoten | yes |
| [4.] Quelle:Tim/Wiedenmann 2000 |
| Autor | Jörg Wiedenmann, Carsten Elke, Klaus-Dieter Spindler, and Werner Funke |
| Titel | Cracks in the b-can: Fluorescent proteins from Anemonia sulcata (Anthozoa, Actinaria) |
| Zeitschrift | PNAS |
| Jahr | 2000 |
| Seiten | 14091–14096 |
| URL | http://www.ncbi.nlm.nih.gov/pmc/articles/PMC18876/ |
| Fußnoten | yes |
| [5.] Quelle:Tim/Wikipedia Snakelocks anemone 2012 |
| Autor | Wikipedia |
| Titel | Snakelocks anemone |
| Jahr | 2012 |
| URL | http://en.wikipedia.org/w/index.php?title=Snakelocks_anemone&oldid=500993724 |
Literaturverz. |
no |
| Fußnoten | no |
| [6.] Quelle:Tim/Lukyanov 2000 |
| Autor | Konstantin A. Lukyanov, Arkady F. Fradkov, Nadya G. Gurskaya, Mikhail V. Matz, Yulii A. Labas, Aleksandr P. Savitskyi, Mikhail L. Markelov, Andrey G. Zaraisky, Xiaoning Zhao, Yu Fang |
| Titel | Natural Animal Coloration Can Be Determined by a Nonfluorescent Green Fluorescent Protein Homolog |
| Zeitschrift | THE JOURNAL OF BIOLOGICAL CHEMISTRY |
| Datum | August 2000 |
| Seiten | 25879–25882 |
| URL | http://www.jbc.org/content/275/34/25879.full.pdf |
Literaturverz. |
yes |
| Fußnoten | yes |
| [7.] Quelle:Tim/Chapleau 2008 |
| Autor | Richard R. Chapleau, Rebecca Blomberg, Peter C. Ford, Martin Sagermann |
| Titel | Design of a highly specific and noninvasive biosensor suitable for real-time in vivo imaging of mercury (II) uptake |
| Zeitschrift | Protein Science |
| Jahr | 2008 |
| Seiten | 614–622 |
| URL | http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2271171/pdf/614.pdf |
Literaturverz. |
yes |
| Fußnoten | yes |
| [8.] Quelle:Tim/Fu 2008 |
| Autor | Yi Fu, Jian Zhang, and Joseph R. Lakowicz |
| Titel | Metal-Enhanced Fluorescence of Single Green Fluorescent Protein (GFP) |
| Zeitschrift | Biochem Biophys Res Commun. |
| Datum | 28. November 2008 |
| Seiten | 712–717 |
| DOI | 10.1016/j.bbrc.2008.09.062. |
| URL | http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2613175/ |
Literaturverz. |
yes |
| Fußnoten | yes |
| [9.] Quelle:Tim/Gurskaya 2003 |
| Autor | Nadya G. GURSKAYA, Arkady F. FRADKOV, Natalia I. POUNKOVA, Dmitry B. STAROVEROV, Maria E. BULINA, Yurii G. YANUSHEVICH, Yulii A. LABAS, Sergey LUKYANOV and Konstantin A. LUKYANOV |
| Titel | A colourless green fluorescent protein homologue from the non-fluorescent
hydromedusa Aequorea coerulescens and its fluorescent mutants |
| Zeitschrift | Biochem. J. |
| Jahr | 2003 |
| Seiten | 403–408 |
| URL | http://www.inbi.ras.ru/competition/2003/articles/labas1.pdf |
Literaturverz. |
yes |
| Fußnoten | yes |
| [10.] Quelle:Tim/Isarankura 2009 |
| Autor | Chartchalerm Isarankura-Na-Ayudhya & Tanawut Tantimongcolwat & Hans-Joachim Galla & Virapong Prachayasittikul |
| Titel | Fluorescent Protein-Based Optical Biosensor for Copper Ion Quantitation |
| Zeitschrift | Biol Trace Elem Res |
| Jahr | 2010 |
| Seiten | 352–363 |
| Anmerkung | online 2009 |
| URL | http://www.pubfacts.com/detail/19649570/Fluorescent-protein-based-optical-biosensor-for-copper-ion-quantitation. |
Literaturverz. |
yes |
| Fußnoten | yes |
| [11.] Quelle:Tim/Sumner et al 2006 |
| Autor | James P. Sumner, Nissa M. Westerberg, Andrea K. Stoddard, Tamiika K. Hurst, Michele Cramer, Richard B. Thompson, Carol A. Fierke, Raoul Kopelman |
| Titel | DsRed as a highly sensitive, selective, and reversible fluorescence-based biosensor for both Cu+ and Cu2+ ions |
| Zeitschrift | Biosensors and Bioelectronics |
| Verlag | Elsevier |
| Ausgabe | 21 |
| Jahr | 2006 |
| Seiten | 1302-1308 |
| DOI | 10.1016/j.bios.2005.04.023 |
| URL | http://www.sciencedirect.com/science/article/pii/S0956566305001752 |
Literaturverz. |
yes |
| Fußnoten | yes |
| [12.] Quelle:Tim/Rahimi et al 2008 |
| Autor | Yasmeen Rahimi, Ann Goulding, Suresh Shrestha, Sweetie Mirpuri, Sapna K. Deo |
| Titel | Mechanism of Copper Induced Fluorescence Quenching of Red Fluorescent Protein, DsRed |
| Zeitschrift | Biochemical and Biophysical Research Communications |
| Ausgabe | 370 |
| Datum | May 2008 |
| Nummer | 1 |
| Seiten | 57-61 |
| Anmerkung | The linked file has page numbers 1-13. Those are used for the documentation. |
| DOI | 10.1016/j.bbrc.2008.03.034 |
| URL | http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2527065/pdf/nihms49229.pdf |
Literaturverz. |
yes |
| Fußnoten | yes |
| [13.] Quelle:Tim/Copper 2010 |
| Titel | Copper |
| Herausgeber | Mark McClure |
| Jahr | 2010 |
| Anmerkung | Part of the "Interactive Periodic Table" http://www2.uncp.edu/home/mcclurem/ptable/ptable.htm |
| URL | http://web.archive.org/web/20100817063208/http://www.uncp.edu/home/mcclurem/ptable/copper/cu.htm |
Literaturverz. |
no |
| Fußnoten | no |
| [14.] Quelle:Tim/MSIP 2010 |
| Titel | Marine Species Identification Portal |
| Datum | November 2010 |
| URL | https://web.archive.org/web/20101130195433/http://species-identification.org/species.php?species_group=zsao&id=2199 |
Literaturverz. |
no |
| Fußnoten | no |
| [15.] Quelle:Tim/Schaefer 2007 |
| Autor | Lars V. Schäfer / Gerrit Groenhof / Astrid R. Klingen / G. Matthias Ullmann / Martial Boggio-Pasqua / Michael A. Robb / Helmut Grubmüller |
| Titel | Photoswitching of the Fluorescent Protein asFP595: Mechanism, Proton Pathways, and Absorption Spectra |
| Zeitschrift | Angew. Chem. |
| Jahr | 2007 |
| Seiten | 536 –542 |
| DOI | 10.1002/ange.200602315 |
Literaturverz. |
yes |
| Fußnoten | yes |
| [16.] Quelle:Tim/Tasdemir et al 2008 |
| Autor | Aynur Tasdemir, Farid Khan, Thomas A. Jowitt, Lucia Iuzzolino, Stefan Lohmer, Sabrina Corazza, Thomas J. Schmidt |
| Titel | Engineering of a monomeric fluorescent protein AsGFP499 and its applications in a dual translocation and transcription assay |
| Zeitschrift | Protein Engineering, Design & Selection |
| Ausgabe | 21 |
| Jahr | 2008 |
| Nummer | 10 |
| Seiten | 613-622 |
| DOI | 10.1093/protein/gzn040 |
| URL | http://peds.oxfordjournals.org/content/21/10/613.full.pdf |
Literaturverz. |
yes |
| Fußnoten | yes |
| [17.] Quelle:Tim/Andresen et al 2005 |
| Autor | Martin Andresen, Markus C. Wahl, André C. Stiel, Frauke Gräter, Lars V. Schäfer, Simon Trowitzsch, Gert Weber, Christian Eggeling, Helmut Grubmüller, Stefan W. Hell, Stefan Jakobs |
| Titel | Structure and mechanism of the reversible photoswitch of a fluorescent protein |
| Zeitschrift | Proceedings of the National Academy of Sciences of the United States of America |
| Ausgabe | 102 |
| Jahr | 2005 |
| Nummer | 37 |
| Seiten | 13070-13074 |
| DOI | 10.1073/pnas.0502772102 |
| URL | http://www.pnas.org/content/102/37/13070.full.pdf |
Literaturverz. |
yes |
| Fußnoten | yes |
| [18.] Quelle:Tim/Schaefer 2007b |
| Autor | Lars Schäfer |
| Titel | Photoactivated Processes in Condensed Phase studied by Molecular Dynamics Simulations |
| Jahr | 2007 |
| Anmerkung | Dissertation University of Braunschweig |
| URL | http://www.mpibpc.mpg.de/278056/Schafer-Lars_2007_Phd-thesis.pdf |
Literaturverz. |
no |
| Fußnoten | no |
| [19.] Quelle:Tim/Wikipedia Conica (suborder) 2009 |
| Titel | Conica (suborder) |
| Verlag | (Wikipedia) |
| Datum | 29. November 2009 |
| Anmerkung | The image in question was "created: May 22, 2008" |
| URL | http://en.wikipedia.org/w/index.php?title=Conica_(suborder)&oldid=328672470 |
Literaturverz. |
no |
| Fußnoten | no |