Angaben zur Quelle [Bearbeiten]
| Autor | Nicole Hellwig |
| Titel | Protonenleitfähigkeit von TRPV1 und Multimerisierung von TRPV-Kanaluntereinheiten |
| Ort | Berlin |
| Jahr | 2005 |
| Anmerkung | Inaugural-Dissertation zur Erlangung des akademischen Grades Dr. rer. nat. am Fachbereich Biologie, Chemie, Pharmazie der Freien Universität Berlin |
| URL | http://www.diss.fu-berlin.de/diss/servlets/MCRFileNodeServlet/FUDISS_derivate_000000001663/ |
Literaturverz. |
nein |
| Fußnoten | nein |
| Fragmente | 8 |
| [1.] Anh/Fragment 006 01 - Diskussion Zuletzt bearbeitet: 2014-09-27 21:01:38 Schumann | Anh, Fragment, Gesichtet, Hellwig 2005, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 6, Zeilen: 1ff (komplett) |
Quelle: Hellwig 2005 Seite(n): 13, Zeilen: 1ff |
|---|---|

Tabelle 1: Regulationsmechanismen und Expressionsorte der TRPC- und TRPV-Kanäle DAG: Diacylglycerol; 5’,6’-EET: 5’,6’-Epoxyeicosatetraensäure; ICRAC: „calcium release-activated calcium current”; InsP3R: Inositol-1,4,5-trisphosphat-Rezeptor; PIP2: Phosphoinsositol-4,5 biphosphat; ROC: rezeptorgesteuert („receptor-operated channel“); SOC: speichervermittelt reguliert („store-operated channels“); WF: Wachstumsfaktor. |

Tabelle EI: Regulationsmechanismen und Expressionsorte der TRPC- und TRPV-Kanäle DAG: Diacylglycerol; 5’,6’-EET: 5’,6’-Epoxyeicosatetraensäure; ICRAC: „calcium release-activatd (sic) calcium current”; InsP3R: Inositol-1,4,5-trisphosphat-Rezeptor; PIP2: Phosphoinsositol-4,5-bisphosphat; ROC: rezeptorgesteuert („receptor-operated channel“); SOC: speichervermittelt reguliert („store-operated channels“); WF: Wachstumsfaktor. |
Ein Verweis auf die Quelle fehlt. Offenbar wurde die Tabelle via copy-paste übernommen. |
|
| [2.] Anh/Fragment 007 17 - Diskussion Zuletzt bearbeitet: 2014-09-27 21:03:42 Schumann | Anh, Fragment, Gesichtet, Hellwig 2005, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 7, Zeilen: 17-31 |
Quelle: Hellwig 2005 Seite(n): 16, Zeilen: 13-27 |
|---|---|
| Transient receptor potential Canonical (TRPC)-Subfamilie
Innerhalb der TRPC-Subfamilie sind 7 Säugerhomologe, TRPC1 bis TRPC7, kloniert worden. Die TRPC-Kanäle sind nicht-selektive, Ca²+-permeable Kationenkanäle und werden nach Stimulation von G-Protein-gekoppelten Rezeptoren oder Rezeptortyrosinkinasen aktiviert („receptor-operated channel“; ROC), was sie somit zu molekularen Korrelaten des rezeptorvermittelten Ca²+-Einstroms macht. Die zugrundeliegenden direkten Aktivierungsmechanismen werden zur Zeit noch sehr kontrovers diskutiert. Dabei scheint es nicht unerheblich zu sein, dass von allen TRPC-Isoformen, außer bei TRPC5, Spleißvarianten existieren, die sich meist durch Deletionen im N- oder C-Terminus unterscheiden. Für TRPC1 konnte bislang kein eindeutiger Aktivierungsmechanismus nachgewiesen werden. Erste Arbeiten deuteten auf eine speicherabhängige Aktivierung hin (Zitt et al., 1996), die jedoch von anderen Arbeitsgruppen nicht bestätigt werden konnte (Sinkins et al., 1998; Brereton et al., 2000). In Studien, bei der die Antisense-Strategie genutzt wurde, konnte jedoch eine Reduktion des durch Speicherentleerung ausgelösten Ca²+-Einstroms gezeigt [werden (Liu et al., 2000; Wu et al., 2000; Brough et al., 2001).] |
TRPC-Subfamilie
Innerhalb der TRPC-Subfamilie sind 7 Säugerhomologe, TRPC1 bis TRPC7, kloniert worden. Die TRPC-Kanäle sind nicht-selektive, Ca2+-permeable Kationenkanäle und werden nach Stimulation von G-Protein-gekoppelten Rezeptoren oder Rezeptortyrosinkinasen aktiviert („receptor-operated channel“; ROC), was sie somit zu molekularen Korrelaten des rezeptorvermittelten Ca2+-Einstroms macht. Die zugrundeliegenden direkten Aktivierungsmechanismen werden zur Zeit noch sehr kontrovers diskutiert. Dabei scheint es nicht unerheblich zu sein, dass von allen TRPC-Isoformen, außer bei TRPC5, Spleißvarianten existieren, die sich meist durch Deletionen im N- oder C-Terminus unterscheiden. Für TRPC1 konnte konnte bislang kein eindeutiger Aktivierungsmechanismus nachgewiesen werden. Erste Arbeiten deuteten auf eine speicherabhängige Aktivierung hin (Zitt et al., 1996), die jedoch von anderen Arbeitsgruppen nicht bestätigt werden konnte (Sinkins et al., 1998; Brereton et al., 2000). In Studien, bei der die Antisense-Strategie genutzt wurde, konnte jedoch eine Reduktion des durch Speicherentleerung ausgelösten Ca2+-Einstroms gezeigt werden (Liu et al., 2000; Wu et al., 2000; Brough et al., 2001). |
Ein Verweis auf die Quelle fehlt. |
|
| [3.] Anh/Fragment 008 01 - Diskussion Zuletzt bearbeitet: 2014-09-27 21:08:05 Schumann | Anh, Fragment, Gesichtet, Hellwig 2005, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 8, Zeilen: 1ff (komplett) |
Quelle: Hellwig 2005 Seite(n): 16, 17, Zeilen: 16: letzte Zeilen; 17: 1ff |
|---|---|
| In der Arbeit von Hofmann et al. (2002) lag TRPC1 jedoch im heterologen Expressionssystem retiniert vor und konnte lediglich durch Coexpression mit TRPC4 bzw. TRPC5 an die Plasmamembran gesteuert werden. In HEK293- Zellen [sic] konnten heterooligomere Kanalkomplexe aus TRPC1 und TRPC5 durch Gq-koppelnde Rezeptoren, nicht aber durch Entleerung intrazellulärer Ca²+- Speicher aktiviert werden, wobei sich TRPC1/TRPC5-Heteromere durch eigenständige biophysikalische Eigenschaften auszeichnen (Strübing et al., 2001). Eine funktionelle Bedeutung könnten TRPC1/TRPC5- Kanalkomplexe im ZNS besitzen, da in Hirnhomogenaten der Ratte durch Immunpräzipitation entsprechende Heteromere nachgewiesen werden konnten (Strübing et al., 2001; Goel et al., 2002). Weiterhin konnte gezeigt werden, dass TRPC1 bei der Sekretion in den β-Zellen des Pankreas beteiligt ist (Sakura und Ashcroft, 1997) und in Purkinje Zellen des Cerebellums sowie in dopaminergen Neuronen nach Aktivierung von metabotropen Glutamatrezeptoren als wichtiger Modulator der synaptischen Aktivität fungiert (Bengtson et al., 2004; Kim et al., 2003; Tozzi et al., 2003).
TRPC2 liegt im Menschen und beim Rind als Pseudogen vor (Wes et al., 1995; Wissenbach et al., 1998). Bei Nagetieren konnte jedoch für TRPC2 eine Expression im Vomeronasalorgan (VNO) und eine Bedeutung bei der Pheromonperzeption nachgewiesen werden (Liman et al., 1999; Leypold et al., 2002). Das Ausschalten des TRPC2-Gens in Mäusen führte bei ihrem Paarungsverhalten zu einem Verlust der Geschlechtsspezifität (Stowers et al., 2002; Keverne, 2002). Es konnte weiterhin gezeigt werden, dass TRPC2 in vomeronasalen Neuronen durch Diacylglycerole aktiviert wird (Lucas et al., 2003). Weiterhin wurde TRPC2 eine Rolle in Spermien bei der Eizellbefruchtung zugewiesen (Jungnickel et al., 2001), jedoch konnte bei Mäusen keine Expression von TRPC2 in Testis nachgewiesen werden und die Fertilität in TRPC2-defizienten Tieren war unverändert (Stowers et al., 2002). Für TRPC2 wurde sowohl eine Aktivierung durch Rezeptorstimulation als auch durch Speichererntleerung gezeigt (Chu et al., 2002; Vannier et al., 1999). In verschiedenen heterologen Expressionssystemen konnte hingegen nur eine intrazelluläre Retention von TRPC2 und keine signifikante Funktion nachgewiesen werden (Hofmann et al., 2000). Die subzelluläre Zielsteuerung von nativ exprimiertem TRPC2 in VNO-Neuronen der Ratte erfolgte jedoch korrekt und zwar in den sensorischen Zilien (Liman et al., 1999). In den sensorischen Mikrovilli des VNO sind aber keine internen Ca²+-Speicher vorhanden, so dass hier eine speichervermittelte Aktivierung von TRPC2 ausgeschlossen werden kann. |
In der Arbeit von Hofmann et al. (2002) lag TRPC1 jedoch im heterologen Expressionssystem retiniert vor und konnte lediglich durch Coexpression mit TRPC4 bzw. TRPC5 an die Plasmamembran gesteuert werden. In HEK293-
[Seite 17] Zellen konnten heterooligomere Kanalkomplexe aus TRPC1 und TRPC5 durch Gq-koppelnde Rezeptoren, nicht aber durch Entleerung intrazellulärer Ca2+-Speicher aktiviert werden, wobei sich TRPC1/TRPC5-Heteromere durch eigenständige biophysikalische Eigenschaften auszeichnen (Strübing et al., 2001). Eine funktionelle Bedeutung könnten TRPC1/TRPC5- Kanalkomplexe im ZNS besitzen, da in Hirnhomogenaten der Ratte durch Immunpräzipitation entsprechende Heteromere nachgewiesen werden konnten (Strübing et al., 2001; Goel et al., 2002). Weiterhin konnte gezeigt werden, dass TRPC1 bei der Sekretion in den β-Zellen des Pankreas beteiligt ist (Sakura und Ashcroft, 1997) und in Purkinje Zellen des Cerebellums sowie in dopaminergen Neuronen nach Aktivierung von metabotropen Glutamatrezeptoren als wichtiger Modulator der synaptischen Aktivität fungiert (Bengtson et al., 2004; Kim et al., 2003; Tozzi et al., 2003). TRPC2 liegt im Menschen und beim Rind als Pseudogen vor (Wes et al., 1995; Wissenbach et al., 1998). Bei Nagetieren konnte jedoch für TRPC2 eine Expression im Vomeronasalorgan (VNO) und eine Bedeutung bei der Pheromonperzeption nachgewiesen werden (Liman et al., 1999; Leypold et al., 2002). Das Ausschalten des TRPC2-Gens in Mäusen führte bei ihrem Paarungsverhalten zu einem Verlust der Geschlechtsspezifität (Stowers et al., 2002; Keverne, 2002). Es konnte weiterhin gezeigt werden, dass TRPC2 in vomeronasalen Neuronen durch Diacylglycerole aktiviert wird (Lucas et al., 2003). Weiterhin wurde TRPC2 eine Rolle in Spermien bei der Eizellbefruchtung zugewiesen (Jungnickel et al., 2001), jedoch konnte bei Mäusen keine Expression von TRPC2 in Testis nachgewiesen werden und die Fertilität in TRPC2-defizienten Tieren war unverändert (Stowers et al., 2002). Für TRPC2 wurde sowohl eine Aktivierung durch Rezeptorstimulation als auch durch Speichererntleerung gezeigt (Chu et al., 2002; Vannier et al., 1999). In verschiedenen heterologen Expressionssystemen konnte hingegen nur eine intrazelluläre Retention von TRPC2 und keine signifikante Funktion nachgewiesen werden (Hofmann et al., 2000). Die subzelluläre Zielsteuerung von nativ exprimiertem TRPC2 in VNO-Neuronen der Ratte erfolgte jedoch korrekt und zwar in den sensorischen Zilien (Liman et al., 1999). In den sensorischen Mikrovilli des VNO sind aber keine internen Ca2+-Speicher vorhanden, so dass hier eine speichervermittelte Aktivierung von TRPC2 ausgeschlossen werden kann. |
Ein Verweis auf die Quelle fehlt. Ein Indiz für eine Copy-&-Paste-Übernahme: Die falsche Leerstelle in "In HEK293- Zellen" befindet sich dort, wo in der Quelle ein Seitenumbruch ist. |
|
| [4.] Anh/Fragment 009 01 - Diskussion Zuletzt bearbeitet: 2014-09-27 21:10:19 Schumann | Anh, Fragment, Gesichtet, Hellwig 2005, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 9, Zeilen: 1ff (komplett) |
Quelle: Hellwig 2005 Seite(n): 17, 18, 19, Zeilen: 17: letzte Zeile; 18: 1ff; 19: 1ff |
|---|---|
| TRPC3, TRPC6 und TRPC7 bilden aufgrund ihrer engen strukturellen Verwandtschaft eine eigene Untergruppe und zeigen auch funktionell eine große Übereinstimmung. Alle drei Kanäle werden durch Ni²+, La³+ und Gd³+ blockiert. Erste Arbeiten zu TRPC3 beschrieben einen speicherabhängigen Aktivierungsmechanismus (Zhu et al., 1996; Preuss et al., 1997). Wenig später wurde jedoch eine Stimulation und Ca²+-abhängige Potenzierung von TRPC3 unabhängig von der Entleerung intrazellulärer Speicher gezeigt (Zitt et al., 1997). Darauf folgende Untersuchungen zur Regulation dieser Kanalfamilie bestätigten diesen Befund und ergaben weiterhin, dass TRPC3, TRPC6 und TRPC7 unabhängig von Proteinkinasen C durch Stimulation der PLC und durch Diacylglycerole aktiviert werden (Hofmann et al., 1999; Okada et al., 1999; McKay et al., 2000). Die Mitglieder dieser Kanaluntergruppe werden u.a. in glatter Muskulatur, Gehirn, Herz und Lunge exprimiert (Tabelle 1). Weitere Arbeiten zeigten, dass TRPC6 in glatten Gefäßmuskelzellen wahrscheinlich den nichtselektiven Kationenstrom nach Stimulation des α1-adrenergen Rezeptors vermittelt (Inoue et al., 2001) und in glatten Muskelzellen aus Ratten (A7r5-Zellen) beim rezeptorvermittelten Einstrom von Ca²+ involviert ist (Jung et al., 2002).
TRPC4 und TRPC5 bilden ebenfalls aufgrund struktureller und funktioneller Ähnlichkeiten eine weitere Kanaluntergruppe innerhalb der TRPC-Subfamilie. Die erste funktionelle Charakterisierung von heterolog exprimiertem bovinem TRPC4 ließ auf einen durch Speicherentleerung aktivierbaren Ca²+-selektiven Kationenkanal schließen (Warnat et al., 1999; Philipp et al., 1996; Philipp et al., 2000). Auch für TRPC5 wurde zunächst eine Aktivierbarkeit durch Speicherentleerung beschrieben (Philipp et al., 1998). In anderen Arbeiten konnten jedoch nicht-selektive Kationenströme in TRPC4- oder TRPC5- exprimierenden Zellen nach Stimulation Gq-koppelnder Rezeptoren aufgezeigt werden (Okada et al., 1998; Obukhov und Nowycky, 2002; Schaefer et al., 2000; Schaefer et al., 2002). Eine weitere Studie zeigte bei Expression von humanem TRPC4 in CHO-Zellen konstitutiv aktive Kationenströme (McKay et al., 2000). Mit Hilfe von Knock-out Mäusen konnte gezeigt werden, dass die Aktivierung speichervermittelt regulierter Kanalkomplexe (SOCs) und die endotheliale Relaxation der Blutgefäße in der Abwesenheit von TRPC4 verändert sind (Freichel et al., 2001). |
TRPC3, TRPC6 und TRPC7 bilden aufgrund ihrer engen strukturellen Verwandtschaft eine
[Seite 18] eigene Untergruppe und zeigen auch funktionell eine große Übereinstimmung. Alle drei Kanäle werden durch Ni2+, La3+ und Gd3+ blockiert. Erste Arbeiten zu TRPC3 beschrieben einen speicherabhängigen Aktivierungsmechanismus (Zhu et al., 1996; Preuss et al., 1997). Wenig später wurde jedoch eine Stimulation und Ca2+-abhängige Potenzierung von TRPC3 unabhängig von der Entleerung intrazellulärer Speicher gezeigt (Zitt et al., 1997). Darauf folgende Untersuchungen zur Regulation dieser Kanalfamilie bestätigten diesen Befund und ergaben weiterhin, dass TRPC3, TRPC6 und TRPC7 unabhängig von Proteinkinasen C durch Stimulation der PLC und durch Diacylglycerole aktiviert werden (Hofmann et al., 1999; Okada et al., 1999; McKay et al., 2000). [...] Die Mitglieder dieser Kanaluntergruppe werden u.a. in glatter Muskulatur, Gehirn, Herz und Lunge exprimiert (siehe Tabelle EI [sic]). Weitere Arbeiten zeigten, dass TRPC6 in glatten Gefäßmuskelzellen wahrscheinlich den nichtselektiven Kationenstrom nach Stimulation des α1-adrenergen Rezeptors vermittelt (Inoue et al., 2001) und in glatten Muskelzellen aus Ratten (A7r5-Zellen) beim rezeptorvermittelten Einstrom von Ca2+ involviert ist (Jung et al., 2002). TRPC4 und TRPC5 bilden ebenfalls aufgrund struktureller und funktioneller Ähnlichkeiten eine weitere Kanaluntergruppe innerhalb der TRPC-Subfamilie. Die erste funktionelle Charakterisierung von heterolog exprimiertem bovinem TRPC4 ließ auf einen durch Speicherentleerung aktivierbaren Ca2+-selektiven Kationenkanal schließen (Warnat et al., 1999; Philipp et al., 1996; Philipp et al., 2000). Auch für TRPC5 wurde zunächst eine Aktivierbarkeit durch Speicherentleerung beschrieben (Philipp et al., 1998). In anderen Arbeiten konnten jedoch nicht-selektive Kationenströme in TRPC4- oder TRPC5-exprimierenden Zellen nach Stimulation Gq-koppelnder Rezeptoren aufgezeigt werden (Okada et al., 1998; Obukhov und Nowycky, 2002; [Seite 19] Schaefer et al., 2000; Schaefer et al., 2002). Eine weitere Studie zeigte bei Expression von humanem TRPC4 in CHO-Zellen konstitutiv aktive Kationenströme (McKay et al., 2000). Mit Hilfe von Knock-out Mäusen konnte gezeigt werden, dass die Aktivierung speichervermittelt regulierter Kanalkomplexe (SOCs) und die endotheliale Relaxation der Blutgefäße in der Abwesenheit von TRPC4 verändert sind (Freichel et al., 2001). |
Ein Verweis auf die Quelle fehlt. In der Quelle sollte es "Tabelle E1" anstatt "Tabelle EI" heißen. |
|
| [5.] Anh/Fragment 010 01 - Diskussion Zuletzt bearbeitet: 2014-09-27 21:10:52 Schumann | Anh, Fragment, Gesichtet, Hellwig 2005, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 10, Zeilen: 1ff (komplett) |
Quelle: Hellwig 2005 Seite(n): 11, 12, Zeilen: 11: 16ff.; 12: 1-4, 11-17 |
|---|---|
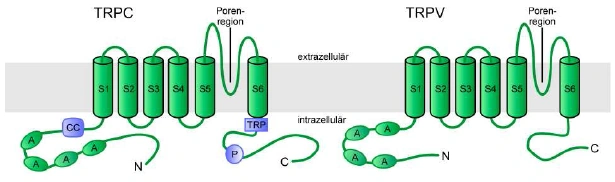
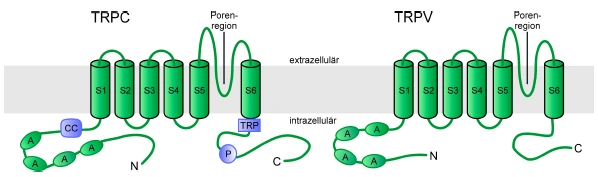
Abbildung 1: Schematische Darstellung der postulierten Domänenstruktur der TRPC- und TRPV Subfamilie Alle TRPC- und TRPV-Kanaluntereinheiten enthalten 6 Transmembransegmente (S1-S6) mit einer putativen Porenregion zwischen dem 5. und 6. Transmembransegment und drei bis vier Ankyrinhomologe Wiederholungen (A). Die Mitglieder der TRPC-Subfamilie besitzen weiterhin eine coiled-coil Domäne (CC) im N-Terminus sowie eine TRP-Box (TRP) und eine prolinreiche Region (P) im CTerminus. Strukturelle Eigenschaften der TRPC Kanäle Alle TRPC- und TRPV-Kanaluntereinheiten besitzen eine Proteinstruktur mit cytosolischen N und C-Termini, sechs putativen Transmembransegmenten und einer putativen Porenregion, welche aus einem kurzen hydrophoben Abschnitt zwischen dem 5. und 6. Transmembransegment besteht (Vannier et al., 1998; Clapham et al., 2003). Die Struktur ähnelt somit jener der spannungsgesteuerten Kaliumkanäle, der durch zyklische Nukleotide regulierten Kanäle (CNG) und der „hyperpolarization-activated and cyclic nucleotide-gated channels” (HCN). Dem 4. Transmembransegment fehlen allerdings die positiv geladenen Aminosäurereste, die in den spannungsgesteuerten Ionenkanälen als Spannungssensor fungieren. Die Kanaluntereinheiten der TRPC- und TRPV-Subfamilien besitzen am N-Terminus mehrere Ankyrin-homologe Wiederholungen („ankyrin-repeats”), deren Funktion innerhalb der TRP-Kanäle noch nicht aufgeklärt ist. Einige der Ankyrin-homologen Wiederholungen können durch Bildung von coiled-coil Strukturen, die Interaktion zwischen TRP-Kanaluntereinheiten und der Ausbildung von Kanalkomplexen vermitteln. In Analogie zu den spannungsgesteuerten 12 Kaliumkanälen und CNG-Kanälen werden funktionelle TRP-Kanalkomplexe durch eine homo bzw. auch heteromere Quartärstruktur aus vier [Kanaluntereinheiten aufgebaut, wie dies bereits für Vertreter der TRPC- und einige Mitglieder der TRPV-Familie gezeigt werden konnte (Kedei et al., 2001; Hofmann et al., 2002; Hoenderop et al., 2003).] |
1.2.1. Strukturelle Eigenschaften
Alle TRPC- und TRPV-Kanaluntereinheiten besitzen eine Proteinstruktur mit cytosolischen N- und C-Termini, sechs putativen Transmembransegmenten und einer putativen Porenregion, welche aus einem kurzen hydrophoben Abschnitt zwischen dem 5. und 6. Transmembransegment besteht (Vannier et al., 1998; Clapham et al., 2003; Abb. E3). Die Struktur ähnelt somit jener der spannungsgesteuerten Kaliumkanäle, der durch zyklische Nukleotide regulierten Kanäle (CNG) und der „hyperpolarization-activated and cyclic nucleotide-gated channels” (HCN). Dem 4. Transmembransegment fehlen allerdings die positiv geladenen Aminosäurereste, die in den spannungsgesteuerten Ionenkanälen als Spannungssensor fungieren. Die Kanaluntereinheiten der TRPC- und TRPV-Subfamilien besitzen am N-Terminus mehrere Ankyrin-homologe Wiederholungen („ankyrin-repeats”), deren Funktion innerhalb der TRP-Kanäle noch nicht aufgeklärt ist. Einige der Ankyrin-homologen Wiederholungen können durch Bildung von coiled-coil Strukturen, die Interaktion zwischen TRP-Kanaluntereinheiten und der Ausbildung von Kanalkomplexen vermitteln. In Analogie zu den spannungsgesteuerten [Seite 12] Kaliumkanälen und CNG-Kanälen werden funktionelle TRP-Kanalkomplexe durch eine homo- bzw. auch heteromere Quartärstruktur aus vier Kanaluntereinheiten aufgebaut, wie dies bereits für Vertreter der TRPC- und einige Mitgleider (sic) der TRPV-Familie gezeigt werden konnte (Kedei et al., 2001; Hofmann et al., 2002; Hoenderop et al., 2003). [...]
Abbildung E3: Schematische Darstellung der postulierten Domänenstruktur der TRPC- und TRPV-Subfamilie Alle TRPC- und TRPV-Kanaluntereinheiten enthalten 6 Transmembransegmente (S1-S6) mit einer putativen Porenregion zwischen dem 5. und 6. Transmembransegment und drei bis vier Ankyrinhomologe Wiederholungen (A). Die Mitglieder der TRPC-Subfamilie besitzen weiterhin eine coiled-coil Domäne (CC) im N-Terminus sowie eine TRP-Box (TRP) und eine prolinreiche Region (P) im CTerminus. |
Ein Verweis auf die Quelle fehlt. |
|
| [6.] Anh/Fragment 011 01 - Diskussion Zuletzt bearbeitet: 2014-09-27 21:12:28 Schumann | Anh, Fragment, Gesichtet, Hellwig 2005, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 11, Zeilen: 1-8 |
Quelle: Hellwig 2005 Seite(n): 11, 12, Zeilen: 11: letzte Zeile; 12: 1-10 |
|---|---|
| [In Analogie zu den spannungsgesteuerten 12 Kaliumkanälen und CNG-Kanälen werden funktionelle TRP-Kanalkomplexe durch eine homo bzw. auch heteromere Quartärstruktur aus vier] Kanaluntereinheiten aufgebaut, wie dies bereits für Vertreter der TRPC- und einige Mitglieder der TRPV-Familie gezeigt werden konnte (Kedei et al., 2001; Hofmann et al., 2002; Hoenderop et al., 2003). Welche Domänen für die Zusammenlagerung erforderlich sind, ist bislang noch unbekannt. Zu den hochkonservierten Regionen innerhalb der TRPC-Subfamilie zählen zudem eine coiled-coil Domäne im cytosolischen N-Terminus, ein kurz hinter dem 6. Transmembransegment befindliches Motiv aus sechs Aminosäuren, das als TRP-Box bzw. nach der Abfolge der Aminosäuren als EWKFAR Motiv bezeichnet wird sowie ein prolinreiches Motiv im zytosolischen C-Terminus. | In Analogie zu den spannungsgesteuerten
[Seite 12] Kaliumkanälen und CNG-Kanälen werden funktionelle TRP-Kanalkomplexe durch eine homo- bzw. auch heteromere Quartärstruktur aus vier Kanaluntereinheiten aufgebaut, wie dies bereits für Vertreter der TRPC- und einige Mitgleider (sic) der TRPV-Familie gezeigt werden konnte (Kedei et al., 2001; Hofmann et al., 2002; Hoenderop et al., 2003). Welche Domänen für die Zusammenlagerung erforderlich sind, ist bislang noch unbekannt. Zu den hochkonservierten Regionen innerhalb der TRPC-Subfamilie zählen zudem eine coiled-coil Domäne im cytosolischen N-Terminus, ein kurz hinter dem 6. Transmembransegment befindliches Motiv aus sechs Aminosäuren, das als TRP-Box bzw. nach der Abfolge der Aminosäuren als EWKFAR-Motiv bezeichnet wird sowie ein prolinreiches Motiv im cytosolischen C-Terminus. |
Ein Verweis auf die Quelle fehlt. |
|
| [7.] Anh/Fragment 012 12 - Diskussion Zuletzt bearbeitet: 2014-09-27 21:14:44 Schumann | Anh, Fragment, Gesichtet, Hellwig 2005, KomplettPlagiat, SMWFragment, Schutzlevel sysop |
|
|
|
| Untersuchte Arbeit: Seite: 12, Zeilen: 12-27 |
Quelle: Hellwig 2005 Seite(n): 5, Zeilen: 2ff |
|---|---|
| Calcium
Das divalente Calciumkation (Ca²+) ist ein ubiquitär in eukaryoten Zellen genutzter intrazellulärer Botenstoff, der diverse physiologische Prozesse kontrolliert. Hierzu gehören u.a. die Ausschüttung von Neurotransmittern, die Kontraktion von Muskelzellen, die Sekretionstätigkeit endo- und exokriner Drüsen, die Gentranskription, die Zellproliferation, die Regulation von Ionenkanälen sowie einer Reihe von Ca²+-abhängigen enzymatischen Stoffwechselprozessen (Ebashi, 1972; Llinás, 1982; Eckert und Chad, 1984; Carafoli und Penniston, 1986; Rasmussen, 1989; Rogers, 1989; Somlyo und Himpens, 1989; Pietrobon et al., 1990; Sheng et al., 1991; Bading et al., 1993; Ghosh und Greenberg; 1995; van Haasteren et al., 1999). Diese zellulären Effekte werden über einen Anstieg der intrazellulären Ca²+-Konzentration ([Ca²+]i) hervorgerufen, der auch als Ca²+-Signal bezeichnet wird. Die internen Ca²+-Quellen befinden sich innerhalb des interzellulären Membransystems, im endoplasmatischen Retikulum (ER), welches bei Muskelzellen als sarcoplasmatisches Retikulum (SR) bezeichnet wird. Im Lumen des ER bzw. SR wird Ca²+ durch Speicherproteine wie Calsequestrin und Calreticulin gepuffert, die eine geringe Ca²+-Affinität jedoch eine hohe Kapazität mit ca. 50 Calciumionen pro Speicherprotein besitzen. |
1.1. Calcium-Homöostase
Das divalente Calciumkation (Ca2+) ist ein ubiquitär in eukaryoten Zellen genutzter intrazellulärer Botenstoff, der diverse physiologische Prozesse kontrolliert. Hierzu gehören u.a. die Ausschüttung von Neurotransmittern, die Kontraktion von Muskelzellen, die Sekretionstätigkeit endo- und exokriner Drüsen, die Gentranskription, die Zellproliferation, die Regulation von Ionenkanälen sowie einer Reihe von Ca2+-abhängigen enzymatischen Stoffwechselprozessen (Ebashi, 1972; Llinás, 1982; Eckert und Chad, 1984; Carafoli und Penniston, 1986; Rasmussen, 1989; Rogers, 1989; Somlyo und Himpens, 1989; Pietrobon et al., 1990; Sheng et al., 1991; Bading et al., 1993; Ghosh und Greenberg; 1995; van Haasteren et al., 1999). Diese zellulären Effekte werden über einen Anstieg der intrazellulären Ca2+-Konzentration ([Ca2+]i) hervorgerufen, der auch als Ca2+-Signal bezeichnet wird. [...] [...] Die internen Ca2+-Quellen befinden sich innerhalb des interzellulären Membransystems, im endoplasmatischen Retikulum (ER), welches bei Muskelzellen als sarcoplasmatisches Retikulum (SR) bezeichnet wird. Im Lumen des ER bzw. SR wird Ca2+ durch Speicherproteine wie Calsequestrin und Calreticulin gepuffert, die eine geringe Ca2+-Affinität (Kd im mM-Bereich) jedoch eine hohe Kapazität mit ca. 50 Calciumionen pro Speicherprotein besitzen. |
Ein Verweis auf die Quelle fehlt. |
|
| [8.] Anh/Fragment 013 01 - Diskussion Zuletzt bearbeitet: 2014-09-27 21:16:43 Schumann | Anh, Fragment, Gesichtet, Hellwig 2005, SMWFragment, Schutzlevel sysop, Verschleierung |
|
|
|
| Untersuchte Arbeit: Seite: 13, Zeilen: 1-10 |
Quelle: Hellwig 2005 Seite(n): 5, 6, 7, Zeilen: 5: 25-28; 6: Bildunterschrift; 7: 8f. |
|---|---|
| [Die] Freisetzung von Ca²+ aus diesen internen Speichern wird durch verschiedene Kanäle kontrolliert, wobei die TRPC Kanäle eine bedeutende Rolle spielen.
In ruhenden Zellen beträgt die [Ca²+]i um 50-100 nM. Nach Stimulation der Zelle kann extrazelluläres Ca²+ über plasmamembranäre Kationenkanäle in das Cytosol gelangen. Hierzu gehören spannungsgesteuerte Ca²+-Kanäle („voltage-operated Ca²+ channels“, VOC), Liganden-gesteuerten Kanäle („ligand-operated channels“, LOC), über sekundäre Botenstoffe gesteuerte Kanäle („second messenger-operated channels“, SMOC) und speichervermittelt regulierten Kanäle („store-operated channels“, SOC). Der Ca²+-Einstrom aus dem extazellulären Medium in das Cytosol wird über Ca²+-permeable Kationenkanäle gesteuert. |
Die Freisetzung von Ca2+ aus diesen internen Speichern wird durch verschiedene Kanäle kontrolliert, wobei die Familie der Inositol-1,4,5-trisphosphat-Rezeptoren (InsP3R) und der Ryanodin- Rezeptoren (RyR) bislang am besten und weitesten untersucht wurden (Abb. E1).
[Seite 6] [...] In ruhenden Zellen beträgt die [Ca2+]i um 50-100 nM. Nach Stimulation der Zelle kann extrazelluläres Ca2+ über plasmamembranäre Kationenkanäle in das Cytosol gelangen. Hierzu gehören spannungsgesteuerte Ca2+-Kanäle („voltage-operated Ca2+ channels“, VOC), Liganden-gesteuerten Kanäle („ligand-operated channels“, LOC), über sekundäre Botenstoffe gesteuerte Kanäle („second messenger-operated channels“, SMOC) und speichervermittelt regulierten Kanäle („store-operated channels“, SOC). [Seite 7] Der Ca2+-Einstrom aus dem extazellulären Medium in das Cytosol wird über Ca2+-permeable Kationenkanäle gesteuert, |
Ein Verweis auf die Quelle fehlt. |
|